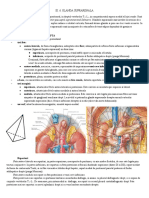
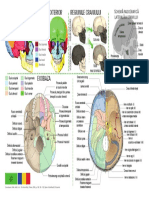
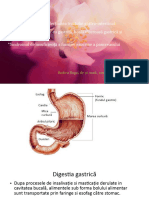
S-ar putea să vă placă și
- Luminita Chiutu Fiziologia Respiratiei Schimburile GazoaseDocument38 paginiLuminita Chiutu Fiziologia Respiratiei Schimburile GazoaseFredutuÎncă nu există evaluări
- Fiziologia RespirațieiDocument10 paginiFiziologia RespirațieiAnonymous CtIkF4vwÎncă nu există evaluări
- New Microsoft Power Point PresentationDocument17 paginiNew Microsoft Power Point Presentationcreatza4uÎncă nu există evaluări
- BiologieDocument20 paginiBiologiePaula CristianaÎncă nu există evaluări
- Personalitati Ale Dezvoltarii Odontoterapiei Restauratoare A Facultatii de Medicina Dentara Din BucurestiDocument6 paginiPersonalitati Ale Dezvoltarii Odontoterapiei Restauratoare A Facultatii de Medicina Dentara Din BucurestimichÎncă nu există evaluări
- Acizi NucleiciDocument12 paginiAcizi NucleiciMihaela Gaidur100% (1)
- Curs 1 Cergan An 2Document92 paginiCurs 1 Cergan An 2GEORGIANA-CIPRIANA CIUBOTARIUÎncă nu există evaluări
- Osteologia Teste Medicina GeneralaDocument21 paginiOsteologia Teste Medicina GeneralanastiuhaÎncă nu există evaluări
- Curs 7 Tesutul Osos PPT PDFDocument45 paginiCurs 7 Tesutul Osos PPT PDFAlex AlxÎncă nu există evaluări
- BiochimieDocument5 paginiBiochimieAlexandra-Maria CostandacheÎncă nu există evaluări
- III.5. Glanda SuprarenalaDocument4 paginiIII.5. Glanda SuprarenalaDiaconu IustinÎncă nu există evaluări
- Fiziologie - Medicina DentaraDocument59 paginiFiziologie - Medicina Dentaragabri1001Încă nu există evaluări
- Combinaţii Heterometalice Ale Cromului (Iii)Document123 paginiCombinaţii Heterometalice Ale Cromului (Iii)mihaelasarateanuÎncă nu există evaluări
- Excre TiaDocument16 paginiExcre TiaStelian CebanasÎncă nu există evaluări
- EchilibrulDocument25 paginiEchilibrulRussu DumitruÎncă nu există evaluări
- Glandele Endocrine - Curs 1Document7 paginiGlandele Endocrine - Curs 1Flavius-Stefan MarinÎncă nu există evaluări
- GRASIMIDocument3 paginiGRASIMIPavel OanaÎncă nu există evaluări
- CURS 12 Compusi DicarboniliciDocument10 paginiCURS 12 Compusi DicarboniliciMerla OanaÎncă nu există evaluări
- Sistem Digestiv CaruntuDocument0 paginiSistem Digestiv CaruntuȘtefania ȘerbănescuÎncă nu există evaluări
- Subiect Simulare 2013 Kineto, Asistenta, Tehnica, Radiologie-Varianta 4Document5 paginiSubiect Simulare 2013 Kineto, Asistenta, Tehnica, Radiologie-Varianta 4Marcella DragutsaÎncă nu există evaluări
- Referat Morfologia Sistemului StomatognatDocument8 paginiReferat Morfologia Sistemului StomatognatCrystyan Crysty Cojocaru100% (1)
- Rolul Rinichilor Si Al Plamanilor in Reglarea Echilibrului Acido-BazicDocument22 paginiRolul Rinichilor Si Al Plamanilor in Reglarea Echilibrului Acido-BazicNiculina CirsteaÎncă nu există evaluări
- Subiecte Pentru Examenul de Anatomie Anul IIDocument4 paginiSubiecte Pentru Examenul de Anatomie Anul IICristina IoanaÎncă nu există evaluări
- Metode de Microscopie S.A.Document30 paginiMetode de Microscopie S.A.Mirela BojogaÎncă nu există evaluări
- Anatomie Sem. 2 - Curs 2Document4 paginiAnatomie Sem. 2 - Curs 2Andreea SchmitzerÎncă nu există evaluări
- Fiziologie Curs - HemodinamicaDocument7 paginiFiziologie Curs - HemodinamicaCiprian DolhescuÎncă nu există evaluări
- Fiziosem 2Document160 paginiFiziosem 2Liliana LupuÎncă nu există evaluări
- Hemodinamica Si Evaluarea Ei-2015 PDFDocument48 paginiHemodinamica Si Evaluarea Ei-2015 PDFIlioaicaMarianaÎncă nu există evaluări
- Raspunsuri La Intrebarile de La Prelegeri Biofizica ExamenDocument4 paginiRaspunsuri La Intrebarile de La Prelegeri Biofizica ExamenpzdtskblÎncă nu există evaluări
- LP 02 BFKTDocument47 paginiLP 02 BFKTGabriel Botezatu67% (3)
- 2 Biochimie Bibl Si SubiecteDocument16 pagini2 Biochimie Bibl Si SubiecteMihaela AntonÎncă nu există evaluări
- Completari SNDocument12 paginiCompletari SNCosmina 2510100% (1)
- Muschii ScaleniDocument2 paginiMuschii ScaleniDaniel Nichi MihaiÎncă nu există evaluări
- Patologie Neurochirurgicala PediatricaDocument93 paginiPatologie Neurochirurgicala PediatricaBogdan ParliteanuÎncă nu există evaluări
- Mecanismul de Funcționare Al Canalelor IoniceDocument4 paginiMecanismul de Funcționare Al Canalelor IoniceEduard ButuÎncă nu există evaluări
- 1echilibrul OsmoticDocument44 pagini1echilibrul OsmoticRachel MNÎncă nu există evaluări
- Biosinteza ARNDocument97 paginiBiosinteza ARNTed Kami-samaÎncă nu există evaluări
- Boron Cap 12Document12 paginiBoron Cap 12miaubooksÎncă nu există evaluări
- Tesuturile Secretoare ExterneDocument21 paginiTesuturile Secretoare ExterneConstantinescu AnamariaÎncă nu există evaluări
- Cifre Corint NetDocument3 paginiCifre Corint NetDary WallÎncă nu există evaluări
- Examene Iarna 2014 StudentiDocument167 paginiExamene Iarna 2014 StudentiCarmen JurcoaneÎncă nu există evaluări
- Sinteza AGDocument71 paginiSinteza AGvictorasaurasÎncă nu există evaluări
- Ru OsukoduDocument1 paginăRu OsukoduRyō Sumy100% (1)
- Sim Dent 2012Document14 paginiSim Dent 2012Roxana BociocÎncă nu există evaluări
- Curs 1Document87 paginiCurs 1Diana LeonÎncă nu există evaluări
- Fenomene de TransferDocument23 paginiFenomene de TransferStancioiuBalaciLauraÎncă nu există evaluări
- CromoproteineleDocument7 paginiCromoproteineleNicolae CudreaÎncă nu există evaluări
- Subiecte Rezolvate Anatomie .An 1 Sem 2Document4 paginiSubiecte Rezolvate Anatomie .An 1 Sem 2Marius FeroiuÎncă nu există evaluări
- Carbonic AnhidrazaDocument7 paginiCarbonic AnhidrazacroitorugeorgianaÎncă nu există evaluări
- Fiziologia Secretiei Salivare.2017Document30 paginiFiziologia Secretiei Salivare.2017Paul EmilianÎncă nu există evaluări
- Fiziologie TotDocument45 paginiFiziologie TotPasvantu DanielÎncă nu există evaluări
- ANATOMIEDocument5 paginiANATOMIE12052000Încă nu există evaluări
- Compusi MacromoleculariDocument28 paginiCompusi MacromoleculariDeni Bv100% (1)
- Respira TiaDocument13 paginiRespira TiaVlad RazvanÎncă nu există evaluări
- FF - Difuzia.osmoza CURS 3Document35 paginiFF - Difuzia.osmoza CURS 3Andra CosmaÎncă nu există evaluări
- Difuzia Si Transportul GazelorDocument6 paginiDifuzia Si Transportul GazelorAdina NeagoeÎncă nu există evaluări
- Subiectul 1Document43 paginiSubiectul 1Bianca AntofieÎncă nu există evaluări
- Resp 3 FiziologieDocument50 paginiResp 3 FiziologieOana Rosca100% (1)
- RespiratiaDocument13 paginiRespiratiagabrielbalan568Încă nu există evaluări
- Preparatele Cu Acţiune Asupra Inervaţiei Periferice Preparatele Colinomimetice Şi AnticolinesteraziceDocument3 paginiPreparatele Cu Acţiune Asupra Inervaţiei Periferice Preparatele Colinomimetice Şi AnticolinesteraziceDaniela DucantoniÎncă nu există evaluări
- Ascultatia Cordului Zgomote, Și SufluriDocument62 paginiAscultatia Cordului Zgomote, Și SufluriCimpoi AureliaÎncă nu există evaluări
- Radiologie Examen Imagini1Document86 paginiRadiologie Examen Imagini1Daniela DucantoniÎncă nu există evaluări
- Farma Lesin AmusDocument2 paginiFarma Lesin AmusDaniela DucantoniÎncă nu există evaluări
- F I Z I o P A T o L o G I A Generală StudiazăDocument1 paginăF I Z I o P A T o L o G I A Generală StudiazăDaniela DucantoniÎncă nu există evaluări
- ANALGEZICE OPIOIDE NEOPIOIDE PT Acad.Document88 paginiANALGEZICE OPIOIDE NEOPIOIDE PT Acad.Daniela DucantoniÎncă nu există evaluări
- Patologia Vasculară. Patologia Cordului. Reumatism. - 0Document6 paginiPatologia Vasculară. Patologia Cordului. Reumatism. - 0Daniela DucantoniÎncă nu există evaluări
- Fiziopatologie, Volumul 1 Din 2Document257 paginiFiziopatologie, Volumul 1 Din 2Doina Haidarli0% (2)
- Ilovepdf MergedDocument44 paginiIlovepdf MergedDaniela DucantoniÎncă nu există evaluări
- TerapieDocument250 paginiTerapieAlexandra ChiricencoÎncă nu există evaluări
- Ar Și Ghini Un 10Document108 paginiAr Și Ghini Un 10Daniela DucantoniÎncă nu există evaluări
- TerapieDocument250 paginiTerapieAlexandra ChiricencoÎncă nu există evaluări
- Corectate La Moment Cu Toate IntrebarileDocument190 paginiCorectate La Moment Cu Toate IntrebarileTatiana JosanÎncă nu există evaluări
- Ilovepdf MergedDocument44 paginiIlovepdf MergedDaniela DucantoniÎncă nu există evaluări
- Teste Romana Semestrul CinciDocument118 paginiTeste Romana Semestrul CinciAnn AnaÎncă nu există evaluări
- Imuno Fiz - Docx 1 1Document42 paginiImuno Fiz - Docx 1 1Marinela BuzanÎncă nu există evaluări
- Ar Și Ghini Un 10Document108 paginiAr Și Ghini Un 10Daniela DucantoniÎncă nu există evaluări
- Fizpat TotalizareDocument38 paginiFizpat TotalizareAudi AudiÎncă nu există evaluări
- Tot 2 FizpatDocument17 paginiTot 2 FizpatDaniela DucantoniÎncă nu există evaluări
- Orarul Sem 5 2023-2024Document1 paginăOrarul Sem 5 2023-2024Daniela DucantoniÎncă nu există evaluări
- 1011 Prelegere - Glucide - Lipide-1Document147 pagini1011 Prelegere - Glucide - Lipide-1LoredanaÎncă nu există evaluări
- AUSCULTATIA PLAMANILOR - STDDocument122 paginiAUSCULTATIA PLAMANILOR - STDDaniela DucantoniÎncă nu există evaluări
- MG III Rom SD Clinice În Gastrită Boală Ulceroasă Pancreatită-87855Document148 paginiMG III Rom SD Clinice În Gastrită Boală Ulceroasă Pancreatită-87855Daniela DucantoniÎncă nu există evaluări
- Exam in Area in Af. A FicatuluiDocument98 paginiExam in Area in Af. A Ficatuluiecaterina_d100% (1)
- Corectate La Moment Cu Toate IntrebarileDocument190 paginiCorectate La Moment Cu Toate IntrebarileTatiana JosanÎncă nu există evaluări
- Teste Morfo 2017 - 5342647233675942243346495053 PDFDocument135 paginiTeste Morfo 2017 - 5342647233675942243346495053 PDFIlie CebanÎncă nu există evaluări
- Farma TabeleDocument2 paginiFarma TabeleDaniela Ducantoni100% (1)
- Combine PDFDocument664 paginiCombine PDFDaniela DucantoniÎncă nu există evaluări
- Bolile FicatuluiDocument3 paginiBolile FicatuluiDora CenusaÎncă nu există evaluări
- Asepsie Rom.Document49 paginiAsepsie Rom.AnaSterpuÎncă nu există evaluări