Documente Academic
Documente Profesional
Documente Cultură
Cavitatea Bucala
Încărcat de
Gigel CovaljievTitlu original
Drepturi de autor
Formate disponibile
Partajați acest document
Partajați sau inserați document
Vi se pare util acest document?
Este necorespunzător acest conținut?
Raportați acest documentDrepturi de autor:
Formate disponibile
Cavitatea Bucala
Încărcat de
Gigel CovaljievDrepturi de autor:
Formate disponibile
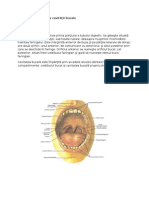
I.CAVITATEA BUCAL Cavitatea bucal este situat n viscerocraniu.
Arcadele alveolo-dentare mpart aceast cavitate n dou poriuni: una antero-lateral, situat n afara arcadelor dentare, denumit vestibul, i alta, nuntrul acestor arcade, denumit cavitate bucal propriu-zis. Aceste dou regiuni comunic ntre ele prin spaiile interdentare i spaiul retrodentar situat napoia ultimilor molari. Anterior, cavitatea bucal comunic prin orificiul bucal cu exteriorul, iar posterior, prin istmul buco-faringian cu faringele. Peretele antero-lateral al cavitii bucale este format din buze i obraji. Mucoasa bucal cptuete pereii cavitii bucale, adaptndu-i structura n fiecare regiune, dup solicitrile din timpul masticaiei, deglutiiei i fonaiei si este perforat de dini. Sistemul de susinere al dinilor este cunoscut sub numele de parodoniu i constituie una din zonele cele mai adaptabile la diverse solicitri funcionale, avnd o sensibilitate crescut, motivat prin bogata vascularizaie i inervaie a acestei regiuni. BUZELE Sunt dou repliuri cutaneo-mucoase, ce delimiteaz prin marginile lor libere orificiul bucal i acoper arcadele alveolo-dentare n partea lor anterioar. Faa anterioar a buzelor este cutanat, fiind prevzut cu foliculi pieloi i glande sebacee, ca i restul tegumentelor faciale. Buza superioar prezint pe linia median un an numit phiftrum, care se lrgete n jos, terminnduse printr-o poriune mai proeminent numit tubercul labiai. De la acest tubercul pleac spre septul nazal o plic median, descris la noi de Rainer. Lateral, buza superioar este desprit de obraz prin anul labiogenian. Buza inferioar este desprit de regiunea mentonier prin anul labiomentonier. Cele dou buze se unesc lateral la nivelul comisurilor bucale. Zona extern, tegumentar a buzelor se continu cu zona intermediar, sau roul buzelor - cunoscut i sub denumirea de zon de tranziie, delimitat de un burelet sau lizereu. Zona de tranziie a buzelor are aceeai origine i structur ca i tegumentele nvecinate, cu deosebirea c roul buzelor este format dintr-un epiteliu stratificat pavimentos nekeratinizat. El prezint un strat lucios dezvoltat i un corion ce conine o bogat reea capilar ce dau culoarea roie acestei regiuni; la rasa alb, n aceast zon, foliculii piloi lipsesc complet, dei glandele sebacee persist nc. La nou-nscut roul buzelor este mprit n dou zone: o zon extern, neted, denumit pars glabra (roul buzelor propriu-zis) i o zon intern, neregulat datorit proeminenelor corionului - pars-villosa, cu rol adjuvantn actul suptului. Zona de tranziie se continu fr linie de demarcaie cu faa intern sau oral a buzei, acoperit de o mucoas de tip bucal care, mpreun cu mucoasa gingival formeaz fundul de sac vestibular sau anul gingivo-labial. Mucoasa labial prezint, ca i tegumentele, un strat epitelial acoperitor i un corion sau derm. Stratul epitelial este format dintr-un strat bazal de celule cilindrice, dintr-un strat malpighian format din celule poliedrice legate ntre ele prin tonofibrile i dintr-un strat superficial de celule cu granulaii de keratohialin. n mod obinuit, aceste celule nu se keratinizeaz dect n cazul respiraiei bucale, datorit uscrii mucoasei de ctre aerul inspirat. Corionul sau dermul mucoasei labiale este bogat n fibre colagene i elastice aici fiind prezente i numeroase glande seroase. O cut vertical a mucoasei, numit frul labial, leag buza n poriunea median gingival. Deseori, la nou-nscui frul labial superior este hiperstratificat, inserndu-se pe mucoasa dintre incisivii centrali superiori ducnd la deprtarea lor (diastem). Scheletul musculo-conjunctiv al buzelor este alctuit din muchiul orbcular al buzelor.
OBRAJII Alctuiesc peretele lateral ai cavitii bucale. Ei sunt formaiuni musculocutanate delimitate anterior de anul nazolabial, posterior de marginea anterioar a muchiului maseter, superior de osul zigomatic i inferior de linia oblic extern a mandibulei. Din punct de vedere structural, obrajii sunt alctuii din: 1. Tegument la exterior - bogat vascularzat i inervat. n corionul tegumentului se afl numeroase glande sudoripare, iar la aduli muli foliculi piloi i glande sebacee. Pe corion se insera i o parte din muchii pieloi ai feei, 2. Stratul celulo-grsos ce conine bula grsoas Bichat, cu important rol n actul suptului, la copil. 3. Stratul muscular format din muchiul buccinator, care-i trimite inserii att pe tegumentele ct i pe mucoasa din aceast regiune. Canalul Stenon al glandei parotide perforeaz aceast fund muscular pentru a se deschide pe mucoasa obrazului, n dreptul celui de-al doilea molar superior. 4. Stratul mucoasei jugale - format dintr-un epiteliu pavimentos stratificat ca i restul mucoasei bucale, ce ader intim la muchiul buccinator situat sub ea, iar n poriunea anterioar mulndu-se pe suprafaa vestibular a dinilor laterali. BOLTA PALATIN Constituie peretele superior al cavitii bucale; ea este format anterior dintr-o poriune osoas rigid palatul dur i posterior dintr-o poriune musculo-aponevrotic supl, mobil - palatul moale sau vlul palatin. Bolta osoas, palatul dur este alctuit anterior apofizele palatine ale oaselor maxilare, iar posterior din lamele orizontale ale oaselor palatine. Pe bolta osoas se afl situate de o parte i de alta a rafeului median, glandele mucoasei palatine. Stratul glandular se oprete la unirea 1/3 anterioare cu cea medie a boltei, aproximativ n dreptul caninilor. Mucoasa palatin ce cptuete palatul dur, se continu anterior i lateral cu mucoasa gingival, iar posterior cu mucoasa vlului palatin. Pe suprafaa ei se distinge pe linia median, rateul palatin, orientat anteroposterior ce corespunde suturii sagitale a palatului osos. La unii indivizi, pe linia median, la nivelul unirii celor dou apofize palatine ale oaselor maxilare se afl cteva proeminene osoase; cunoscute sub numele de torus palatin. n partea anterioar a rafeului median, n dreptul orificiilor incisivilor centrali superiori, mucoasa palatin prezint o proeminen a raeului median, numit papil incisiv. n 2/3 anterioare ale palatului dur, de o parte i de aita a rafeului median, mucoasa palatin prezint 3-7 creste rugoase transversale, oblice sau arciforme, cunoscute sub numele de rugi palatine. La nivelul zonei de separaie dintre palatul dur i palatul moaie, de o parte i de alta a liniei mediane, se afl foveele palatine mici depresiuni n care se deschid canalele excretorii ale glandelor. n 1/3 posterioar, mucoasa palatin este uor mamelonat, datorit glandelor mucoase subiacente. Palatul dur se continu posterior cu palatul moale sau vlul palatin - formaiune musculomembranoas mobil, acoperit de mucoasa vlului palatin care este subire i neted. Linia de demarcaie ntre mucoasa fix a palatului dur i mucoasa mobil a palatului moale, poart numele de linia Ah ce are forma unei linii curbe deschise anterior. Marginea posterioar a vlului palatin este liber i prezint o prelungire median, numit luet. Marginile laterale ale vlului palatin se fixeaz pe pereii laterali ai faringelui. Faa anterioar a vlului palatin prezint un rafeu median, n prelungirea rafeului palatin dur, iar faa posterioar a sa este orientat spre faringe. Vlul palatin se continu pe laturi cu cte dou cute - stlpii vlului palatin, ce delimiteaz lojele amigdaliene n care se afl amigdalele palatine. Datorit muchilor tensori, ridictori, cobortori ai vlului, acesta are un rol important n procesele de deglutiie i fonaie. De asemeni, vlul palatin prezint o bogat vascularizaie i inervaie, fiind sediul a numeroi exteroreceptori responsabili de apariia reflexului de vom sau de deglutiie. PLANEUL CAVITII BUCALE - sau peretele su inferior este un sept muscular situat ntre arcul mandibular i osul hioid, format din unirea pe linia median a celor doi muchi milohioidieni care, alturi de muchii geniohioidieni i pntecele anterior al digastricului, formeaz diafragma bucal.
Se delimiteaz astfel etajul supramilohioidian, n care se afl glandele sublinguale, i etajul submilohioidian, n care se afl glandele submaxiiare. La interior, planeui bucal este acoperit de limb, ce contribuie la delimitarea regiunilor sublingual, lingual i suprahioidian. Regiunea subungual - prezint un repliu mucos median numit frul limbii, care leag faa inferioar a limbii de planeul bucal. Pe laturile frului se afl dou cute transversale ale mucoasei, numite plicile sublinguaie sub care se afl glandele sublinguale care se deschid prin canalele excretoare Rivinius i Bartholini, la nivelul carunculelor sublinguale. De o parte i de alta a frului lingual se afl i carunculele de vrsare a canalului Wharton. Mucoasa sublingual este vascularizat de artera sublingual care se anastomozeaz cu artera submentonier, ramur din facial. Limfaticele zonei dreneaz limfa n ganglionii submaxilari. Fiind subire, bogat vascuiarizat i umed, mucoasa sublingual constituie o membran biologic care permite schimburi ntre mediul intern i mediul bucal. La acest nivel pot fi absorbite substane introduse n cavitatea bucal (alcoolul, unele medicamente). n regiunea lingual se afl limba, organ musculo-fibros, care ocup aproape n ntregime cavitatea bucal propriu-zis. Ea se muleaz pe suprafaa palatin a dinilor, i prezint o poriune posterioar cu mobilitate redus - baza limbii ancorat pe peretele inferior ai cavitii bucale i pe structurile vecine i o poriune interioar mobil - vrful limbii. n structura sa se disting mai multe elemente: fibre musculare striate ce aparin muchilor intrinseci sau extrinseci ai limbii, un schelet fibros pe care se insera fibrele musculare, formaiuni limfoide care constituie amigdala lingual, vase, nervi i mucoasa lingual ce o acoper la exterior. Muchii intrinseci sunt aezai n trei planuri: - Planul superior - conine muchiul lingual superior ce se dispune de la baz la vrful limbii. Contracia fibrelor sale produce scurtarea limbii. - Planul mijlociu - conine fibre musculare dispuse transversal, ntre muchiul lingual superior i lingual inferior. Contracia lor produce ngustarea limbii, conferindu-i forma de jgheab. - Planul inferior - este alctuit din muchii linguali inferiori. Contracia lor produce coborrea i retragerea vrfului limbii. Printre aceste fibre musculare ale planurilor menionate, exist i fibre verticale a cror contracie produce turtirea limbii. Un rol important n mobilizarea limbii n diverse direcii l are i musculatura extrinsec a sa, reprezentat de muchii geniogloi, a cror contracie trage limba nainte i o fixeaz napoia simfizei mentoniere; muchii stilogloi ce trag limba napoi i n sus; muchii palatogloi i faringogloi ce trag rdcina limbii napoi i n sus, aplicnd-o pe bolta palatin. nveliul extern al limbii este reprezentat de mucoas: cea sublingual este subire, elastic, lsnd s se vad prin transparen vasele sanguine i prezentnd median repliul frului limbii ce face legtura cu arcada alveolar inferioar. De o parte i de alta a frului limbii se afl canalele excretoare ale glandelor mucoase descrise de Nuhn-Blandin. Mucoasa feei dorsale a limbii este mai groas i prezint numeroase supradenivelri numite papile linguale sau gustative ce conin mugurii gustativi. Forma, mrimea si dispoziia papilelor gustative este diferit pe suprafaa dorsal a limbii. Astfel, n regiunea V-ului lingual situat la unirea a 2/3 anterioare cu 1/3 posterioar a limbii, se afl 9-11 papile caliciforme sau circumvalate; fiecare papil conine numeroi muguri gustativi i este nconjurat la baz de un an (vallum), n care se deschid glandele salivare seroase von Ebner. Papilele caliciforme sunt cele mai voluminoase, avnd o nlime de 1,5 mm i o grosime de aproximativ 1 mm. Pe marginile laterale ale limbii sunt depuse mai ales papile foliate, n numr redus la om, care sunt formate din 8-10 pliuri mucoase ce pot conine n grosimea epiteliului lor muguri gustativi. Pe faa dorsal a limbii se afl dou tipuri de papile gustative: - papilele filiforme - sunt cele mai numeroase (500/cm2), au o nlime de aproximativ 2,5 mm, au form cilindric sau conic i sunt rspndite pe toat faa dorsal a limbii, naintea "V"-ului lingual i pe marginile ei. Aceste papile sunt acoperite de un epiteliu ce prezint granule de keratohialin. Papilele filiforme nu conin muguri gustativi, avnd rol numai n sensibilitatea tactil; 3
- papilele fungiforme - au o form de ciuperc rotund, sunt rspndite pe toat suprafaa dorsal a limbii; au o nlime de aproximativ 1 mm, sunt foarte bogat vascularizate, ceea ce face s se identifice la suprafaa limbii ca nite puncte mici, roii; n grosimea epiteliului lor de acoperire se afl puini muguri gustativi. La baza limbii, napoia "V"-ului lingual, se afl amigdala lingual ce aparine cercului limfatic Waideyer. Limba este un organ ce conine numeroi receptori: la nivelul mucoasei linguale (receptori gustativi); la nivelul stratului submucos (receptori ai sensibilitii tactile, termice i dureroase) ct i proprioreceptori situai n musculatura intrinsec a limbii, toi acetia furniznd informaii asupra caracteristicilor obiectelor introduse n cavitatea bucal, fiind implicai n reglarea masticaiei, deglutiiei, secreiei salivare, n declanarea reflexelor gustative ct i n vorbirea articulat. MUCOASA BUCAL - sau mucoasa oral, cptuete la interior cavitatea bucal. Histologic, este alctuit dintr-un epiteliu i un corion, separate printr-o membran bazal. Epiteliul, de tip stratificat pavimentos, este alctuit din trei straturi: - stratul bazal germinativ - format din celule prismatice, nalte ce prezint mitoze frecvente, asigurnd rennoirea continu a celulelor superficiale; - stratul spinos pluristratificat - format din celule poliedrice, mai turtite nspre suprafa; - stratul superficial - format din dou-trei rnduri de celule turtite, cu nuclei picnotici, aplatizai, sau fr nuclei, ce se descuameaz continuu. Printre celulele epiteliale ale stratului bazal i spinos se pot gsi: melanocite ce pot sintetiza melanin, conferind pigmentaia caracteristic mucoasei. O intens activitate a melanocitelor ce determin o pigmentaie accentuat a mucoasei bucale, se poate nscrie n sfera patologicului (boala Addison); - celule Merkel ce sunt implicate n procesele senzoriale; celule Langerhans, limfocite, granulocite care dein un rol important n procesele de aprare nespecific. Corionul mucoasei bucale, sau lamina propria este un esut conjunctiv pluricelular bogat n fibre de colagen, macrofage, limfocite, plasmocite, canale excretorii ale glandelor salivare mici, vase sanguine, limfatice i filete nervoase. La nivelul mucoasei obrajilor i mucoasei linguale corionul prezint papilele adelomorfe. Sub corionul mucoasei bucale se afl stratul submucos format din esut conjunctiv lax, acini secretori ai glandelor salivare mici i receptori nervoi senzitivi. Gradul de mobilitate al mucoasei bucale este dat de dezvoltarea stratului submucos. Mucoasa bucal cu mobilitate redus, fix, acoper o parte din bolta palatin i procesele alveolare; are o rezisten sczut la presiune mecanic, lezndu-se uor, i o sensibilitate dureroas accentuat. Mucoasa bucal mobil, cu un strat submucos bine reprezentat acoper buzele, obrajii, vlul palatin, planeul bucal i fundurile de sac de la baza proceselor alveolare. n funcie de caracteristicile structurale, zona de dispunere i rolul su fiziologic, mucoasa bucal se poate mpri n: mucoasa jugal sau a obrajilor; mucoasa labial - pe faa intern a buzelor; mucoasa planeului oral al regiunii sublinguale; mucoasa palatin ce prezint fenomene de keratinizare - este fix n zona vlului palatin; mucoasa lingual - bogat n 2/3 anterioare ale dosului limbii n receptori gustativi; mucoasa gingival (gingia) ce este o mucoas fix cu rol n masticaie (adaptat solicitrilor mecanice). Mucoasa bucal joac un rol important n organism, datorit funciilor sale fiziologice: - funcia de protecie mecanic i de aprare antibacterian; - funcia de resorbie a moleculelor lipo i hidrosolubile (astfel ajungnd n circulaia general unele medicamente); - funcia de excreie a unor substane toxice produse de metabolismul endogen sau a unor bacterii i virusuri; funcia de exteroreceptor pentru sensibilitatea termic, tactil, gustativ. Mucoasa bucal este zon reflexogen pentru reflexele masticatorii, de deglutiie, salivare .a., prin ndeplinirea acestor roiuri fiziologice, mucoasa bucal reprezentnd o membran biologic activ, la nivelul creia pot fi reflectate nenumrate afeciuni patologice generale.
MUCOASA GINGIVAL Acoper procesele alveolare i se continu cu mucoasa vestibular, labial, jugal, sublingual sau palatinal. Ea este perforat de dini. Dup apariia dinilor se pot distinge dou pri ale mucoasei gingivale: gingia liber - rsfrnt spre cementul dentar i gingia aderent - fixat la periostul alveolar. Gingia liber prezint o poriune marginal (bureletul gingival) ce nconjoar coletul dentar i o poriune papilar ce se interpune ntre dini (la btrni aceast zon se retract, se atrofiaz, lsnd spaiul interdentar liber). Bureletul gingival este congestionat n perioada de erupie dentar, apoi i revine, meninndu-i rolul de a mpiedic fragmentele alimentare mari s se insinueze printre dini. ntre bureletul marginal i coroana dintelui se formeaz un fund de sac sub 2 mm (la adult) n care se pot gsi resturi alimentare i se poate depune tartrul. Marginea liber a gingiei se rsfrnge deci pe coletul dentar, adernd ns intim de cementul radicular prin aa numita "inserie epitelial", cu ajutorul tonofibrilelor Baratieri. La nivelul corionului mucoasei gingivale exist numeroase leucocite, datorit microleziunilor produse de coninutul fundului de sac gingival. n cazul unei infecii masive, microbii pot invada mezenchimul subiacent, iar epiteliul gingival prolifereaz spre apex, descriind paradontopatia cronic progresiv. n cazul keratinizrii epiteliului bureletului gingival, datorit unei tulburri metabolice, mucoasa gingival se atrofiaz, celulele epiteliale se decoleaz, inseria epitelial se detaeaz producnd paradontoz. Mucoasa gingival se continu cu restul mucoasei bucale printr-o zon de tranziie, numit mucoasa alveolar, mobil, cu un epiteliu subire, un corion srac n papile i un strat submucos bogat n vase i n esut conjunctiv lax. Mucoasa alveolar este fix numai n regiunea palatinal, unde ader intim la periostul oaselor bolii palatine. PARADONIUL Constituie sistemul de susinere al dintelui i este alctuit din: cement, ligamentul alveolo-dentar, osul alveolar i stratul corial al mucoasei gingivale. Spre deosebire de dinte, care se dezvolt din mugurele dentar, parodoniul se dezvolt din sacul fibros din jurul mugurelul dentar, avnd vascularizaie proprie. Parodoniul are rolul de a transmite presiunile exercitate pe dini asupra pereilor alveolari. Cementul - este asemntor din punct de vedere structural cu esutul osos compact, fiind format din precipitarea srurilor de calciu n ochiurile reelei de esut mezenchimal. Cementul acoper dentina n poriunea radicuiar a dintelui, protejnd-o cnd este lezat, prin apariia de fibre colagene (Sharpey) cu orientri diferite. Ele iau forma unui evantai ce se insera n cement pe de o parte, i n osul alveolar pe de alt parte, intrnd n spaiul desmodontal n alctuirea ligamentului alveolo-dentar. Prin procesele de apoziie i resorbie pe care le sufer, cementul contribuie la modificarea spaiului parodontal. Ligamentul alveolodentar sau desmodontiul - este reprezentat de fascicule de fibre colagene cu orientare diferit n spaiul parodontal (fibre marginale, cemento-papilare, transseptate sau cementocementare, cemento-alveolare). ntre fibrele cemento-alveolare exist un plex fibros (Kerebel) ce permite adaptarea fibrelor colagene la diferite presiuni sau traciuni la care este supus parodoniul n timpul masticaiei. Tot n ochiurile reelei colagene, printre fasciculele fibroase, se afl elemente celulare: fibroblati, fibrocite, celule mezenchimale i numeroase vase cu rol nutritiv. Vasele capilare, ca o adaptare la micrile ample ale coletului dentar n timpul masticaiei i la presiunile exercitate asupra dintelui, prezint dilataii cu rol de amortizare a ocurilor mecanice - denumit de ctre Wesky - "frn hidraulic". n jurul acestor dilataii, exist o bogat reea nervoas vegetativ. Integritatea structural a desmodoniului i normofuncoionalitatea sa sunt deosebit de importante. El asigur ancorarea dintelui n alveolele osoase, controlul micrilor dentare de mic amplitudine i particip la traciunea dintelui n timpul erupiei sale. De asemeni, datorit bogatelor anastomoze vasculare i dublei inervaii (senzitiv i vegetativ), desmodoniul transmite stimulii exteroceptivi (tactili i dureroi) ct i proprioceptivi (ce controleaz micrile de deschidere i nchidere ale gurii n timpul masticaiei) de-a lungul ramurilor nervilor trigemeni i controleaz debitul sanguin al vaselor dentare. 5
Osul dentar sau procesul alveolar mrginete spaiul periodontal i este alctuit din zona cortical extern, subgingival, iar la nivelul alveolei este delimitat de corticala intern, mai dens i perforat, numit lamina dura sau lamina cribriforma. ntre cele dou zone corticale se afl esut osos spongios, ce se continu cu esutul osos al maxilarului propriu-zis. Procesele alveolare creeaz spaiile alveolelor dentare n care se vor afla rdcinile dinilor. ntre peretele alveolar i rdcina dintelui exist spaiul alveolo-dentar de aproximativ 0,1 mm, ocupat de ligamentul alveolo-dentar, vase, nervi i lichid interstiial. Cementul, ligamentul alveolo-dentar, vasele, nervii, lichidul interstiial, procesele alveolare i mucoasa gingival alctuiesc mpreun parodoniul, sistemul de ancorare al dintelui n alveola dentar. II. MORFOFIZIOLOGIA DINILOR I A ARCADELOR DENTRARE STRUCTURA DINILOR PERMANENI Dinii sunt organele cele mai adaptate structural funciei lor, fiind implantai pe cele dou maxilare i formnd arcadele dentare: superioar i inferioar. Omul prezint n cursul vieii dou dentiii: una temporar ("de lapte") i alta permanent (definitiv), ntre vrsta de 6 luni i 2 ani i 6 luni, apar pe rnd cei 20 de dini temporari, iar ntre 6 i 12 ani acetia sunt nlocuii cu cei 32 de dini permaneni. Macroscopic, la nivelul oricrui dinte se disting dou pri structurale: o cavitate central, denumit camera pulpar, n care se afl mnunchiul vasculo-nervos i odontoblatii, i o parte dur, extern, format din dentin. n regiunea coroanei dintelui, dentina este acoperit de un strat mult mai dur, numit smal dentar, iar n regiunea rdcinii dintelui, dentina este acoperit de cement. ntre cement i peretele alveolar se afl un spaiu - alveolo-dentar sau periodontal de 0,10 mm n care se gsete esut conjunctiv bogat n fibre colagene, care suspend dintele ca ntr-un hamac. n timpul procesului de masticaie, presiunile exercitate asupra dinilor constituie fore de traciune asupra peretelui alveolar, stimulnd apoziia osoas i ducnd la apariia unei zone de esut dens pe peretele alveolar, denumit "lamina dura". Pulpa dentar Este un esut conjunctiv situat n zona central a dintelui, n interiorul cavitii pulpare, delimitat la exterior de dentin. Pulpa comunic cu structurile parodontale la nivelul apexului, prin orificiul apical. n compoziia pulpei intr: substana fundamental prin care sunt vehiculate substanele metabolic active necesare vitalitii dintelui; celulele pulpare - care n funcie de morfologia i metabolismul lor se mpart n trei grupe: 1. celule de baz ale esutului pulpar (fibroblati) ce elaboreaz precursorii colagenului i elementele substanei fundamentale. Odat cu naintarea n vrst, fibroblatii sunt nlocuii cu fibrocite - mai srace n organite celulare, cu activitate metabolic redus; 2. celule dentinogenetice - reprezentate de odontoblati i celulele zonei subdonto-blastice. Odontoblatii sunt situai la periferia pulpei coronare i radiculare. Ei elaboreaz componentele matricei dentinare. Celulele din zona subdontoblastic -celulele Hhl sau celulele rotunde Weil, sunt cele care n condiii reacionale (traumatice, infecioase sau terapeutice) au activitate dentino-genetic, putndu-se diferenia n odontoblati ct i n fibroblati. ntre odontoblati i celulele Hohl, se afl zona celular Weil, n care se gsesc capilare ale reelei vasculare subodontoblastice, terminaii nervoase, plexul fibrelor Retterer format din fibrele Korff. 3. celule de aprare (histiocite, pfasmocite, monocite) cu rol antiinflamator i antiinfecios. Histiocitele i macrofagele sunt importante pentru fagocitoza resturilor tisulare i a particulelor strine; plasmocitele, derivate din limfocite au rol n sinteza anticorpilor n procesele inflamatorii ale pulpei dentare; mastocitele bogate n histamin i heparin intervin n mecanismele antialergice i antihemoragice. 6
Apariia unor procese inflamatorii poate induce o stimulare a activitii celulelor pulpare, cu precdere a odontoblatilor, ducnd la formarea unei dentine reacionale. La acest proces contribuie i celulele subodontoblastice ce se pot diferenia n neoodontoblati. Pe de alt parte, n cursul proceselor inflamatorii cronice, prin stimularea osteoclastelor pot aprea zone de reabsorbie ale dentinei, perturbndu-se metabolismul celulelor pulpare. Fibrele pulpare sunt reprezentate de: 1. fibrele colagene, cele mai numeroase, sunt elaborate de fibroblati. Odat cu mbtrnirea, fibrele de colagen se nmulesc i devin tot mai dense. 2. fibrele de reticulin care formeaz n pulpa dintelui o reea n jurul pereilor vasculari. n spaiul Weil (subodontoblastic) se afl fibrele Korff care sunt formate din unirea mai multor fibre de reticulin, care trec printre odontoblati i ajung n dentin, unde se transform n fibre de colagen. 3. fibrele oxitalanice (preelastice) sunt mai puin numeroase dect fibrele de colagen i cele de reticulin, sunt rspndite n toat pulpa dentar i sunt singurele care nu sufer transformri n timp. Vascularizaia pulpei dentare Pulpa prezint o bogat reea vascular de tip terminal, condiionnd vitalitatea dintelui prin asigurarea aportului de substane necesare desfurrii proceselor metabolice la acest nivel. Capilarele pulpare sunt cele la nivelul crora se realizeaz schimburile ntre snge i esutul pulpar. n jurul capilarelor (mai bine reprezentate n zona subodonto-blastic) se pot gsi fibre de reticulin, care protejeaz peretele vascular i permit vasului s se adapteze variaiilor debitului sanguin. Modificarea debitului sanguin prin creterea presiunii arteriale sau prin spasm refiex, determin dilatarea sistemului capilar i stimularea filtrrii plasmatice la nivelul peretelui capilar. Permeabilitatea capilar crescut, duce la apariia stazei, i edemului pulpar cu modificarea consecutiv a proprietilor fizico-chimice ale substanei fundamentale. Datorit spaiului rigid n care se afl pulpa dentar, edemul su poate provoca ruperea endoteliului vascular cu apariia microhemoragiilor pulpare, compresia structurilor vecine i alterrii ireversibile ale esutului pulpar. Vasele limfatice prezint anastomoze ntre limfaticele pulpei dentare i cele ale spaiului parodontal. Drenajul limfatic se face ctre ganglionii submentonieri i submaxilari i apoi ganglionii cervicali. Datorit limfocitelor prezente n ganglioni, vascularizaia limfatic are un rol important n procesele antiinflamatorii i antiinfecioase. Invervaia pulpei dentare Este reprezentat de fibre senzitive ce aparin ramurilor maxilar i mandibular ale nervului trigemen. Ele ptrund, mpreun cu vasele sanguine i fibrele vegetative, prin orificiul apical al rdcinii, fiind fibre mielinice. La nivelul pulpei coronare, fibrele senzitive se ramific n evantai, formnd n zona subodontobiastic plexul Rashkow. La acest nivel i pierd teaca de mielin i trimit terminaii libere printre odontoblati. Fibrele vegetative, simpatice i parasimpatice, nsoesc fibrele senzitive, producnd vasoconstricie (aciune simpatic -adrenergic) sau vasodilataie (aciune parasimpatic -colinergic), n funcie de perturbrile locale. Variai factori fizici (temperatur, presiune) sau chimici, dac depesc o anumit intensitate (prag) pot provoca durerea. Sensibilitatea dureroas prezint variaii, i depinde probabil de numrul de terminaii nervoase pe milimetru ptrat (ntre 30 i 300/mm2). esutul pulpar, bogat n celule, fibre, vase, terminaii nervoase, sufer n timp, odat cu vrsta modificri caracteristice: fibroblatii sunt nlocuii treptat cu fibrocite; odontoblatii involueaz i scad numeric; n spaiul intercelular apar vacuole mari pline de lichid; esutui pulpar se densific i fibrozeaz, peretele vaselor sanguine sufer o distrofie hiperplazic ce duce la ngustarea lumenului vascular; apar osteoblati sau cementoblati care stimuleaz depunerile de sruri minerale i apariia calcificrilor pulpare. Acestea din urm sunt formaiuni de form i mrime diferite, numite pulpolii sau denticuli. Volumul pulpar diminueaz ca urmare a apoziiei continue de dentin. Toate aceste modificri duc la ncetinirea proceselor metabolice i reducerea capacitii de aprare a pulpei dentare. Dentina
Este un esut dur ce nfoar camera pulpar i este acoperit la exterior de smal (la nivelul coroanei dintelui) i de cement (la nivelul rdcinii dentare). Din punct de vedere structural, se disting o dentin manta (periferic) i o dentin circumpulpar (ortodentina). Dentina manta este subire, avnd o grosime de 80-100 micromicroni, este atubular i prezint o strom organic format din fibrele colagene von Korff. Dentina circumpulpar conine un sistem de tubuli sau canalicule dentinare n form de "S" care se deschid n pulpa dintelui. Direcia acestor tubuli este diferit: sunt paraleli cu axul lung al dintelui, n dentina coronar ce corespunde feei ocluzale ale dintelui; sunt oblici ctre feele laterale ale dintelui, iar la nivelul rdcinii sunt perpendiculari pe axul lung al dintelui. Numrul canaliculelor dentinare variaz ntre 20.000 i 65.000/mm2. Ei sunt nconjurai de dentina peritubular, intertubular, prelungirile citoplasmatice ale odontoblatilor (fibrele Tomes cu rol secretor), formnd mpreun unitatea metabolic dentinar. Din punct de vedere al compoziiei, dentina conine 10-15% ap, 60-65% substane anorganice i 30-35% substane organice. Componentul mineral cel mai important este reprezentat de cristalele de hidroxiapatit. Odat cu vrsta, n raport cu solicitrile fiziologice sau factorii patologici, dentina sufer permanent remanieri. Cele mai importante procese de remaniere sunt apoziia dentinar i resorbia dentinar. Apoziia dentinar fiziologic are loc n tot cursul vieii, procesul de formare al dentinei prin mineralizarea matricii elaborate de odontoblati, fiind nentrerupt. Dentinogeneza este stimulat ele asemeni i n condiii patologice (existenta unei spine iritative, de exemplu a cariilor dentare). Aceast dentin este neregulat i imperfect structurat. Resorbia dentinar poate fi i ea un proces fiziologic sau patologic. Resorbia fiziologic se produce prin stimularea osteoclastelor la nivelul rdcinii dinilor temporari i faciliteaz nlocuirea lor cu dentiia definitiv. Reabsorbia dentinar patologic este generat de procese inflamatorii pulpare, de compresie radicular prelungit, de procese infecioase pulpare sau parodontale, cu localizri la diferite segmente ale dintelui. Datorit existenei terminaiilor nervoase dentinare, provenite din pulpa dentar, dentina este sensibil la durere mai ales n absena protejrii ei de ctre smal. Soluii hiperconcentrate acre sau dulci, diferene mari de temperatur ale alimentelor introduse n cavitatea bucal, procesele inflamatorii pulpare, agresiunile mecanice externe pot genera senzaia de durere dentar. Sensibilitatea dureroas dentinar este variabil de la un individ la altul datorit repartiiei inegale a fibrelor nervoase intradentinare, provenite din fibrele pulpare. Smalul Acoper dentina n regiunea coroanei dentare, fiind un esut dur, cu un coninut bogat de sruri minerale (fosfat de calciu i magneziu, clorur de sodiu i potasiu). Fiind un esut acelular, el nu are posibilitatea de a se reface n cazul lezrii sale. Smalul este format din prismele adamantine ce conin cristale de hidroxiapatit i substana interprismatic. Prismele sunt n numr de 5-12 milioane/dinte, fiind dispuse paralel cu axul lung al dintelui n regiunea superficial a coroanei, apoi oblic pn la jonciunea sa cu cementul, iar la acest nivel sunt perpendiculare pe axul lung al dintelui. Substana interprismatic conine o reea de fibrile de natur proteic n ochiurile creia se gsesc proteine solubile i cristale de hidroxiapatit cu diferite orientri. La suprafaa smalului prismatic exist un strat aprismatic numit cuticula smalului cu o grosime de aproximativ 1 micron. Procesele inflamatorii sau nfecioase dentare, intoxicaiile cu fluor sau excesul de tetracicline pot produce defecte de mineralizare ale smalului, reflectate n modificri macroscopice ale sale. De asemeni, contactul prelungit cu soluii acide (scderea pH-ulul n cavitatea bucal) la cei cu igien bucal deficitar, poate produce n timp distrucia smalului prin demineralizarea sa i apariia cariei dentare. Cementul Intr n alctuirea sistemului de susinere al dintelui, prezentat anterior n cadrul parodoniului. El acoper dentina n poriunea rdcinii dintelui, pe faa sa extern inserndu-se ligamentul desmodontal (alveolo-dentar). Ca i structur, se aseamn esutului osos, dar, spre deosebire de acesta, este lipsit de vase i nervi, motiv pentru care are posibiliti reduse de remaniere. Comparativ cu dentina i smalul, cementul conine cea mai redus cantitate de substane anorganice, duritatea sa fiind de 1,4 ori mai mic dect a 8
dentinei i de 2,4 ori mai mic dect a smalului. Cel mai important constituent anorganic al su, fosfatul de calciu, se afl sub form de cristale de hidroxiapatit de dimensiuni foarte mici. De o importan major este i fluorul care, la nivelul cementului, are cea mai mare concentraie comparativ cu dentina i smalul. n mod fiziologic, odat cu vrsta, procesul de mineralizare al cementului se intensific, crescnd concentraia cristalelor de hidroxiapatit i a fluorului. Din punct de vedere structural exist un cement primar (fibrilar sau acelular) care se depune primul deasupra dentinei radiculare, n cursul odontogenezei, n special n regiunea cervical a rdcinii i un cement celular. Acesta din urm acoper cementul primar, iar n regiunea apical a rdcinii acoper direct dentina. Cementul celular conine celule (cementoblaste i cementocite, situate n spaii lacunare numite cementoplaste), fibre de colagen i substan fundamental ce conine sruri de calciu. n mod normal, cementul este continuat la nivelul coletului dintelui de ctre smal. Jonciunea smal-cement nu las n mod fiziologic dentina descoperit, denudarea ei fiind ntlnit n rare cazuri i constituie un loc predilect pentru apariia cariei i o zon cu sensibilitate dureroas crescut. Procesele de remaniere ale cementului, dei reduse, pot fi fiziologice (mbtrnirea de exemplu, stimuleaz apoziia cementar mai ales la nivelul apexului dentar). Prin apoziia fiziologic a cementului este pstrat integritatea structurii i funciei aparatului ligamentar periodontal. Patologic, n afeciunile periapicaie i parodontale poate aprea apoziia cementar reacional. Cea de-a doua modalitate de remaniere a cementului o constituie reabsorbia cementar. Ea se produce fiziologic n cursul rizalizei dinilor temporari, prin stimularea osteoclastelor. Patologic, n afeciuni parodontale, dar i n exercitarea prelungit a compresiei ocluzale, poate aprea resorbia cementar reactiv, urmat de mobilizarea dintelui. ASPECTUL GENERAL AL DINILOR n general, la orice dinte se distruge o poriune vizibil, coroana, cu cinci fee: faa mezial spre linia mediosagital; faa distal sau lateral; faa palatinal sau lingual - n raport cu limba; faa vestibuiar sau labial - n raport cu vestibulul cavitii i faa ocluzal sau triturant n contact cu dinii arcadei opuse, ce realizeaz tierea i zdobirea alimentelor. O alt poriune distinct a dintelui este rdcina, care este implantat n alveola dentar i care se termin cu un vrf sau apex ce prezint un orificiu prin care ptrunde mnunchiul vasculo-nervos dentar. O alt zon, coletul sau gtul dintelui este poriunea acoperit de mucoasa gingival, fiind locul unde smalul se continu cu cementul dintelui. n procesul de masticaie, implicarea dinilor este diferit, ceea ce explic i marea varietate a formei lor. Incisivii - sunt dinii cu rol important n apucarea i tierea hranei. Cei 4 incisivi superiori, au form de lopat, iar cei 4 inferiori, form de dalt. Cei mai mari sunt incisivii centrali superiori, urmnd n ordine: incisivii laterali superiori, incisivii laterali inferiori i incisivii centrali inferiori. Incisivii centrali superiori prezint pe marginea lor ocluzal 3 zimi vizibili din momentul erupiei. Pe faa palatinai a acestor dini exist un tubercul mamelonat, denumit cingulum. Cei 3 zimi i cingulum-ul reprezint de fapt cei 4 centri odontogeni de cretere ai dintelui. Cingulum-ul delimiteaz cu faa palatinal o foset (foramen caecum dentis) deseori locul de placare al cariei. Rdcina incisivilor centrali superiori este unic, avnd form conic, n momentul erupiei, aceti dini au apexul larg deschis, nchizndu-se abia dup 4 ani de la momentul erupiei. Spre deosebire de incisivii centrali superiori, cei centrali inferiori sunt mai mici i nu prezint foramen caecum. Incisivii laterali superiori sunt situai de o parte i de alta a incisivilor centrali superiori, au rdcina mai turtit meziodistal, ca de altfel toi ceilali dini, iar apexul lor este deviat distal, nchzndu-se tot la 4 ani dup erupie. Spre deosebire de acetia, incisivii laterali inferiori sunt mai mari dect incisivii centrali inferiori. Incisivii dentiiei temporare au structur similar incisivilor permaneni, dar se caracterizeaz printr-o camer pulpar mai larg. Caninii - sunt cei mai puternici dini monoradiculari, fiind folosii la prinderea i sfierea hranei. La om, caninii au form de pumnal, cu o coroan pentagonal. Pe faa palatinat (superior) sau lingual (inferior) se disting cele 3 creste ce converg spre coletul dentar ntr-un tubercul mameionat (cingulum). 9
Rdcina caninilor este unic, lung i turtit lateral. Apexul su se nchide n jurul vrstei de 15 ani i este deviat distal. Caninii inferiori sunt mai lungi dect cei superiori; smalul lor coboar mai jos pe faa vestibular dect pe faa lor lingual. Caninii temporari au structur similar, dar o camer pulpar i un canal radicular mai largi. La om, incisivii i caninii fac parte din grupul dinilor frontali. Pe lng rolul lor estetic, acest grup de dini sunt importani n masticaie, n ghidarea micrilor de propulsie i lateralitate ale mandibulei. n ghidajul micrilor laterale ale mandibulei, caninul are rolul principal, mai ales caninul superior, fiind dintele capabil s suporte singur ntregul ghidaj. Totodat, grupul dinilor frontali, prin contactele pe care le stabilesc cu dinii vecini, transmit presiunile masticatorii pe ntreaga arcad, evitndu-se fenomenul de suprasolicitare mecanic a unui singur dinte. Premolarii sunt dispui cte 4 pe fiecare maxilar, ntre canini i molari. Primii premolari superiori (unul drept i unul stng) au o coroan turtit meziodistal; pe faa ocluzal prezint un cuspid vestibular mai voluminos i altui palatina! mai mic. Rdcina primului premolar este divizat n dou rdcini, cea vestibular fiind mai voluminoas. n majoritatea cazurilor au 2 canale radicuare. Al doilea premolar superior se situeaz de fiecare parte ntre primul premolar i primul molar superior. Ei au coroana mai mic; pe faa ocluzal prezint doi cuspizi egali ca volum (unul vestibular i altul palatinal). Rdcina lor este turtit mezio-distal fr a fi divizat. Camera pulpar este turtit meziodistal i prezint prelungiri spre cei doi cuspizi. Canalul radicular este de obicei unic. Premolarii inferiori sunt mai mici dect cei superiori, i au pe faa ocluza doi cuspizi mai puin proemineni (unul vestibular i altul iingua). Pe fiecare cuspid se pot distinge cte 3 creste ce se prelungesc pe feele premolarului. Rdcina premolarilor inferiori este unic, conic, turtit mezio-distal, ca i camera pupar, apexul este deviat iar canalul radicuJar este de obicei unic. Molarii - sunt dinii care au suferit cele mat multe modificri n seria animal, n funcie de tipul alimentaiei. Molarii sunt voluminoi, au form cuboid i un numr mare de cuspizi i rdcini. Primul molar superior este cunoscut sub numele de molarul de 6 ani, deoarece apare la aceast vrst. Coroana sa are form cuboid, cu 5 fee, faa ocluzal prezentnd 3-4 cuspizi, separai printr-un an n forma literei "H" - loc de elecie pentru caria dentar. anul intercuspidian se continu pe faa vestibular a molarului (formnd 2 lobi conveci), ct i pe faa sa palatinal, mai mic, mprind-o i pe aceasta n 2 lobi. Primul molar superior are 3 rdcini divergente, 2 vestibulare i una palatinal, mai puternic. Camera pulpar prezint cte o prelungire ctre fiecare cuspid, iar rdcina prezint 3 canale radiculare. Al 2-lea molar superior, sau molarul de 12 ani, este mai mic dect primul molar superior, prezint pe faa ocluzal de obicei 3 cuspizi (2 vestibulari i unul palatinal). Molarul al 2-lea superior, are tot 3 rdcini i 3 canale radiculare, uneori cele 2 rdcini vestibuiare se unesc, fr a se uni ns i canalele radiculare. Spre deosebire de acesta, al 2-lea molar superior temporar prezint pe faa ocluzal 4 cuspizi i 3 rdcini divergente, asemnndu-se cu primul molar superior permanent. Al 3-lea molar superior, sau molarul de minte, prezint cele mai variate forme i dimensiuni. Poate avea 3 cuspizi ca i al 2-lea molar, dar uneori poate avea i un singur cuspid i o singur rdcin. De obicei, cele 3-4 rdcini ale acestui molar sunt sudate, dar pot fi i divergente, ngreunnd extracia sa. Camera pulpar este larg i se continu cu 3 canale radiculare ce pot fi unite n unul singur. Molarii inferiori sunt cei mai voluminoi dini, cu numrul cel mai mare de cuspizi. Primul molar inferior, molarul de 6 ani are 5 fee, pe faa ocluzal prezint 5 cuspizi (3 vestibulari i 2 linguali). Faa vestibular este convex, fr bose, dar prezint un an vertical, numit "an de despovrare", alimentele triturate scurgndu-se pe acest an. Primul molar inferior are dou rdcini, cea mezial mai voluminoas, deviat distal. Pn la 14 ani, rdcina mezial are un singur canal radicular, iar dup aceast vrst, 2 canale radiculare meziale (unul vestibular i altul lingual). Camera pulpar are forma coroanei i prezint prelungiri spre cuspizi. Primul molar inferior temporar prezint pe faa ocluzal numai 4 cuspizi. Al 2-lea molar inferior este cunoscut ca i molarul de 12 ani, are dimensiuni mai mici dect primul molar inferior; pe faa ocluzal prezint 4 cuspizi, iar feele vestibular i lngual divizate prin anuri 10
verticale. Rdcinile acestui molar sunt puin voluminoase ce au tendina de a se apropia. n unele cazuri, rdcina mezial prezint un singur canal radicular chiar i dup vrsta de 14 ani. n dentiia temporar, al doilea molar inferior este asemntor primului molar inferior permanent, dar cu o suprafa ocluzal mai redus. Al 3-lea molar inferior sau molarul de minte este pe cale de dispariie. Faa ocluzal prezint de obicei ntre 1 i 4 cuspizi, iar rdcinile lor sunt n general unite; cnd sunt divergente n poriunea apical, face extracia lor foarte dificil i cateterizarea canalelor radicutare aproape imposibil. Premoiarii i molarii alctuiesc grupul dinilor laterali. Spre deosebire de dinii frontali ce realizeaz contacte pe o suprafa restrns, suprafaa lor ocluzal fiind ngust, adaptat pentru tierea alimentelor, dinii grupului lateral realizeaz un contact interdentar mult mai larg. Aceasta asigur contacte stabile datorit cuspizilor prezeni pe suprafeele ocluzale. Aceti dini sunt perfect adaptai pentru zdrobirea i mcinarea alimentelor. Aceste procese sunt realizate prin micri de lateralitate ale molarilor ce au cuspizi mai mici, dar aceste micri au efect abraziv al dinilor ct i micri verticale ce predomin la dinii cu cuspizi mai mari, cum sunt premoiarii. Acest tip de micri masticatorii, verticale, pstreaz se pare, mai mult timp, integr suprafaa ocluzal a dinilor, avnd un efect abraziv mult mai redus. n ceea ce privete dimensiunea dinilor, aceasta este n medie mai redus la dinii temporari fa de cei permaneni. n medie, suma diametrelor mezio-distale ale dinilor temporari este cu 2,8 mm mai mic dect suma acelorai diametre ale dinilor permaneni. Rezult c, pn la apariia molarilor de 6 ani, arcada alveolar a dinilor temporari trebuie s creasc n medie cu cel puin 2,8 mm n plan sagital pentru a putea cuprinde toi dinii permaneni. Culoarea dinilor, albicioas albstrie la dinii temporari difer de culoarea galben-gri a dinilor permaneni, aceasta explicndu-se prin diferentele structurale. Astfel, grosimea smalului i a dentinei este mai redus la dinii temporari cu pn la 50%. UNITATEA FUNCIONAL A ARCADEI DENTARE Cele dou arcade dentare se formeaz n urma erupiei dinilor ntr-o anumit succesiune la nivelul celor doua maxilare. Cele dou erupii dentare (temporar i definitiv), respectiv cele dou dentiii, se deosebesc att pnn numrul, morfologia dinilor ct i prin raporturile ocluzale cu dinii antagoniti ai arcadei opuse. Numrul dinilor (10 pe fiecare arcad n dentiia temporar: 4 incisivi, 2 canini i 4 molari); 16 pe fiecare arcada n dentiia permanent (4 incisivi, 2 canini 4 premolan i 6 molari) ct i forma i dimensiunile cuspizilor, a crestelor i fosetelor, sunt determinate genetic. Prin migrarea dinilor n timpul erupiei se stabilete raportul dintre suprafaa dentar superioar i cea inferioar, cunoscut sub numele de ocluzie. ERUPIA DENTAR Erupia dinilor temporari se realizeaz ntre vrsta de 6 luni i 2 ani i jumtate, respectnd urmtoarea secven: 1 - incisivii centrali mandibulari; 2 - incisivii centrali maxilari; 3 - incisivii laterali maxilari; 4 incisivii laterali manclibulari; 5 - primii molari mandibulari i maxilari; 6 - caninii mandibulari i maxilari; 7 - molani secunzi mandibulari; 8 - molarii secunzi maxilari n general, dinii mandibulari erup naintea celor maxilari, cu excepia incisivilor laterali care erup ntro secven invers. Exist numeroi factori fiziologici care influeneaz erupia dinilor temporari, dintre care: greutatea la natere (copiii cu greutate mare la natere prezint apariia timpurie a incisivilor, ntre 3 i 5 luni); sexul copilului (incisivii apar mai devreme la biei; dar molarii i caninii mai repede Ia fete); vrsta mamei i numrul de nateri (erupia este precoce la copii mamelor tinere i este ntrziat la cei ai mamelor multipare); momentul naterii (naterile premature se nsoesc de ntrzieri n erupia primilor dini). n mod patologic, rahitismul, sifilisul congenital i rubeola prenatal se nsoesc de ntrzieri n erupia dinilor temporari; de asemeni, n sindromul Down, ntrzierea erupiei dentare se poate nsoi de o secven 11
neobinuit (erupia primului molar naintea incisivului lateral); febra este un alt factor care accelereaz erupia dentar. Erupia dinilor permaneni se desfoar dup o pauz de civa ani, ntre 6 i 13 ani, cu excepia molarilor de minte care apar mult mai trziu, n genera! dup 16 ani. Secvena erupiei dinilor permaneni este urmtoarea: 1 - primii molari inferiori; 2 - primii molari superiori; 3 - incisivii centrali inferiori 4 - incisivii centrali superiori; 5 - incisivii laterali inferiori; 6 - incisivii laterali superiori; 7 - primii premolari superiori la baiei 8 - caninii inferiori 7 - caninii inferiori la fete 8 - primii premoiari superiori 9 - primii premolari inferiori; 10 - premolarii secunzi inferiori; 11 - premoiarii secunzi superiori; 12 - molarii secunzi inferiori; 13 - molani secunzi superiori. Se observ c dinii mandibulei erup naintea ceior maxilari, cu excepia primilor premolari, la care erupia de la arcada superioar se face naintea celei de la mandibul. Se observ de asemeni c, ntre cele dou sexe exist diferene n ordinea n care erup caninul inferior i primul premolar superior. Factorii fiziologici care influeneaz momentul apariiei dinilor permaneni sunt urmtorii: sexul (la fete, dinii permaneni apar cu 3-5 luni mai devreme); rasa (cea mai rapid erupie dentar o au copii de ras neagr, urmeaz apoi rasa alb; indienii prezint erupia ntrziat a primilor molari i a incisivilor centrali, dar i o erupie mult mai accelerat a celorlali dini, comparativ cu rasa alb); clima tropical accelereaz erupia dentar comparativ cu cea temperat; factorii soclo-economici (copii ce au condiii precare de trai prezint o erupie ntrziat); urbanismul (copii din ora au erupii dentare timpurii comparativ cu cei de la sate); momentul erupiei de-a lungul anilor (n populaia actual, molarii secunzi, caninii i primii pramolari mandibulari apar mai devreme dect n trecut, iar perioada total a erupiei dinilor permaneni s-a scurtat n mod evident de-a lungul anilor). n mod patologic, hipersecreia hormonului de cretere i a hormonilor tiroidieni produc accelerarea erupiei dentare, n timp ce bolile genetice (sindromul Down, Turner) ntrzie apariia dinilor permaneni alturi de defectele nutriionale i metabolice. De asemeni, angioamele feei, hipervasculare, atrag dup ele i accelerarea erupiei dentare; pierderea precoce a dinilor temporari ca urmare a proceselor infecioase dentare conduce la erupia precoce a dinilor permaneni; deplasarea dinilor vecini, cu limitarea spaiului de erupie ca urmare a pierderii premature a dintelui temporar, duce la ntrzierea dintelui permanent. O erupie ntrziat a unui dinte poate fi determinat i de dezvoltarea insuficient a procesului alveolar nsoit de reducerea creterii rdcinii dintelui. Alturi de dini, fibrele muchilor aparatului dento-maxilar se adapteaz prin contraciile i relaxrile lor la tipul micrilor mandibulei, buzelor, limbii, obrajilor, contribuind la realizarea actului masticaiei, deglutiiei, respiraiei, mimicii. Impulsurile proprioceptive culese de la baroreceptorii din spaiile alveolo-dentare i din zonele de inserie muscular, realizeaz prin arcuri reflex condiionate, tipuri de aciuni musculare diferite, care persist chiar i dup cderea sau erodarea dinilor. Nepotrivirea dintre forma i dimensiunile cuspizilor dinilor antagoniti, ct i influena altor factori generali sau locali ce determin o dezvoltare neadecvat a dinilor, a proceselor alveolare sau a maxilarelor genereaz disfuncionaliti ntre cele dou arcade dentare i un tip anormal de aciune muscular. 12
Forma dinilor, adaptat la condiiile de alimentaie, rmne unul dintre elementele cele mai conservatoare din structura organismului uman. Pe de alt parte, forma maxilarelor i tipul de aciune a muchilor sunt n principal date de forma i relaiile dintre dini, constituind un tot funcional. De asemeni, un sistem unitar l constituie i articulaia dentar mpreun cu sindezmozele alveolo-dentare, sindezmozele cranio-faciale i diartroza temporomandibular. III. SECREIA SALIVAR GLANDELE SALIVARE Saliva reprezint un amestec de secreii produse de glandele salivare principale i accesorii, la care se adaug lichidul crevicular sau gingival. Glandele salivare principale sunt glandele parotide, submaxilare i sublinguale. Ele sunt situate n grosimea pereilor cavitii bucale, n lojile corespunztoare. Secreia lor este de tip merocrin, iar structura, tubulo-acinoas, are ca unitate morfofuncional salivonul, format dintr-un acin glandular i canalul sau excretor. Mai muli acini formeaz un lobul care prezint irigaie sanguin i limfatic proprie i este drenat de un canal unic ce rezult din reeaua de canale mici din interiorul lobulului. Glanda parotid - se afl n regiunea parotidian limitat anterior de marginea posterioar a mandibulei, superior de conductul auditiv extern, posterior de mastoid i muchiul sternocleidomastoidian, iar inferior, de o linie convenional ce continu marginea inferioar a mandibulei pn la ntlnirea cu muchiul sternocleidomastoidian. Este cea mai voluminoas gland salivar i are o greutate de 25-50 grame. nvelit n fascia parotidian, glanda trimite 2 prelungiri, o prelungire faringian i o prelungire anterioar ce trece peste suprafaa muchiului maseter i se continu cu poriunea extraglandular a canalului excretor a lui Stenon. Canalul parotidian trece deasupra maseterului, strbate muchiul buccinator i se deschide n vestibulul cavitii bucale, n anul gingivo-alveolar superior, n dreptul rdcinii celui de-al doilea molar superior. Glandele parotide au parenchimul format din acini glandulari i canale excretoare, din care rezult canalul unic - Stenon. Pentru c acinii sunt exclusiv de tip seros, ele sunt considerate glande seroase al cror produs de secreie fluid, transparent, bogat n enzime, reprezint saliva de diluie. Fiecare acin este format din 2-6 celule seroase, aflate pe o membran bazal. Celulele acinare prezint n regiunea bazal un nucleu sferic i un numr moderat de ribozomi, iar apical aparatul lui Golgi i numeroase granule secretorii. Membrana luminal prezint numeroase prelungiri mici, iar pereii laterali, zone de pliere n care jonciunile dintre dou celule alturate devin sinuoase. Spre poriunea apical, uneori i spre cea bazal jonciunea celular este foarte strns. ntre celulele seroase i membrana bazal exist celule stelate mioepiteliale ce prezint elemente contractile care contribuie la evacuarea secreiei primare. Poriunea iniial a sistemului de ducte - ductul intercalat - este delimitat de un epiteliul scuamos cu celule mici cuboidale i deseori celule mioepiteliale plate, cu prelungiri scurte, i are rol n transportul salivei, Duetele intercalate dreneaz n duetele striate, mai mari, delimitate de un epiteliu simplu cilindric cu celule nalte, poligonale i o membran bazal cu numeroase pliuri Pliurile membranei din partea bazal a celulei ptrund adnc n citoplasm, separnd-o n compartimente ce conin mitocondrii dispuse paralel. Acest tip de specializare observat i la nivelul tubului contort distal al nefronului este caracteristic epiteliilor implicate n schimburile rapide de ap i ioni. Citoplasm apical conine numeroase vezicule i granule de diverse mrimi i densiti, sugernd participarea acestor celule la procese de secreie. Duetele intralobulare, intercalate i striate, sunt drenate n duetele interlobulare, mrginite de un epiteliu mai nti columnar apoi pseudostratificat. Canalul principal, n apropierea orificiului de deschidere este limitat de un epiteliu scuamos stratificat. Produsul de secreie al celulelor seroase se caracterizeaz printr-o important activitate amilazic, este bogat n proteine i polizaharide (sialomucin, sulfomucin). Glandele submaxilare (submandibulare) se afl sub planeul bucal ntr-un spaiu aflat ntre mandibul i muchiul digastric, iar n profunzime, se ntind pn la muchii milohioidieni, hipoglos i 13
stiloglos. Canalul lor excretor - canalul Wharton - se vars n vrful carunculei linguale, uneori unindu-se ntr-un confluent comun cu ductul excretor principal al glandei sublinguale. Deoarece n structura lor intr muli acini seroi dar i acini mucoi i micti, submaxilarele sunt glande mixte. Celulele seroase conin granule secretorii PAS-pozitive, bogate n sialoglicoproteine. Celulele mucoase conin fie sialomucin, fie sulfomucin, fie ambele. Sistemul de ducte este similar celui din parotid, dar prezint ducte intercalare mai scurte i ducte striate mai bine reprezentate. n ductele mari, epiteliul pseudostratificat modific compoziia salivei prin secreia de mucus a celulelor i secreia unui fluid seros, realizat de celulele principale columnare. Saliva produs de aceste glande se caracterizeaz printr-o slab activitate amilazic i prin prezena lizozimului secretat de celulele seroase semilunare; este o saliv mai vscoas, cu mai mult mucin numit i saliva de gustaie. Glandele sublinguale sunt situate deasupra planeului bucal, sub mucoasa membranoas de o parte i de alta a liniei mediane, n lojele sublinguale, care adpostesc pe lng glandele sublinguale, prelungirea glandei submandibulare, canalul lui Wharton, nervii lingual i hipoglos, artera lingual i vase limfatice. Glanda sublingual este nvelit ntr-o lam fibroconjunctiv subire i ocup cea mai mare parte a spaiului sublingual. Ea const dintr-o aglomerare de mici glande care toate la un loc au lungimea de 25-30 mm, nlimea 11-15 mm i grosimea 5-6 mm avnd o greutate de 6-8 grame. Produsul de secreie al acestor glande se vars ntr-un canal principal - canalul Bartholin care se deschide lng canalul Wharton sau mpreun cu acesta de o parte i de alta a liniei mediane i n 5-6 canale mici accesorii - canalele Rivinius, care se deschid de o parte i de alta a liniei mediane de-a lungul plicii sublinguale. Glandele sublinguale sunt de tip mucos, avnd majoritatea acinilor formai din celule mucoase, uneori remarcndu-se i prezena unor celule seroase semilunare. Celulele mucoase au forma i aspectul celor seroase, dar prezint un reticul endoplasmatic mai slab dezvoltat i granule secretorii mari, cu aspect vacuolar. Coninutul principal al secreiei de tip mucos este dat de polizaharidele sulfatate. Rarele uniti seroase prezente, conin glicoproteine sulfatate. Acinilor mucoi li se asociaz celule mioepiteliale. Ductele intercalare i striate sunt scurte i mai puin evidente. Capsula glandei este mai ngust i prezint mai puine septuri. Glandele sublinguale secret o saliv bogata n mucus - saliva de deglutiie, cu rol n formarea bolului alimentar i alunecarea acesteia n faringe i esofag n timpul deglutiiei. Glandele salivare accesorii sunt rspndite n stratul submucos din toat mucoasa bucal, fiind mai abundente i grupate n anumite zone: grupul glandelor palatine, n mucoasa vlului palatin; grupul glandelor labiojugale n mucoasa buzelor i a obrajilor; grupul glandelor linguale n mucoasa lingual. Vascularizaia glandelor salivare este abundent, realiznd un debit sanguin local foarte mare (0,3 ml/g esut/minut pentru glandele nestimulate) de 20 de ori mai mare fa de debitul muchiului n repaus. Prin stimulare parasimpatic debitul crete de aproximativ 5 ori. Glandele parotide sunt irigate prin ramuri arteriale provenite din artera facial i carotid extern. Glandele submandibulare primesc ramuri glandulare din artera facial i carotid extern. Glandele sublinguale sunt irigate din artera sublingual i submental. Vasele se distribuie n glande, paralel cu ductele principale inter i intralobulare, alturi de trunchiurile nervoase. Deplasarea sngelui n vase se face n contracurent cu saliva din duete. Exist o capilarizare iniial la nivelul duetelor, arteriolele se refac i apoi se recapilarizeaz la nivelul acinilor. n acest fel se realizeaz circulaia contracurent necesar formrii salivei prin procese de secreie acinar, reabsorbie i secreie tubular. Rezistena acinar crescut la nivelul polului eferent realizat de capilarele periacinare este important pentru procesul de ultrafiltrare acinar. Sngele venos este colectat n vena jugular, iar limfaticele urmeaz traiectul canalelor excretoare, ajungnd la ganglionii limfatici regionali. Inervaia glandelor salivare este realizat de sistemul nervos vegetativ simpatic i parasimpatic. Neuronii simpatici preganglionari se afl la nivelul segmentelor toracale T1 i T2. Fibrele lor preganglionare se distribuie neuronilor simpatici postganglionari din ganglionii cervicali superiori. Fibrele postganglionare se distribuie glandelor salivare. Neuronii parasimpatici preganglionari se afl n nucleii salivatori superior i inferior. Fibrele lor preganglionare sunt incluse n nervii cranieni facial i glosofaringian i se conecteaz cu neuronii parasimpatici postganglionari ai ganglionilor vegetativi din vecintatea glandelor. Fibrele 14
postganglionare scurte ptrund n glande, terminaiile nervoase amielinice simpatice i parasimpatice distribuindu-se celulelor acinare, cte 5-10 axoni pentru fiecare celul acinar. SALIVA - PROPRIETI, COMPOZIIE, ROL Produsul de secreie al celor trei perechi de glande salivare mari i a multiplelor glande salivare mici, diseminate n mucoasa bucal se caracterizeaz prin proprieti i compoziie ce variaz n limite destul de largi n funcie de subiect, natura stimulului aplicat, debitul secreiei etc. Aceste variaii sunt realizate pe de o parte prin modificarea contribuiei cantitative a celor trei secreii, parotidian - seroas, submaxilar mixt, sublingual mucoas, i pe de alt parte prin modificarea calitativ a fiecrui tip de secreie. Glandele salivare ale omului secret n 24 de ore, n medie 1000-1500 (2000) ml saliv. n condiii de repaus, strict standardizate, debitul salivei mixte a fost determinat la 0,35 - 0,5 ml/min, cu variaii de la un individ la altul. n condiii de stimulare reflex, debitul crete la 1,5-2,3ml/min, iar prin stimulare cu droguri parasimpaticomimetice, la 5 ml/min. Din totalul secreiei bazale, submaxilarele contribuie cu 70%, parotidele cu 25%, sublingualele cu 4%, glandele diseminate cu 1%. Dublarea debitului secretor modific nivelul de participare pentru parotid la 34% iar pentru cele submaxilare la 63%, n somn, debitul scade foarte mult, pn la 0,05ml/min, parotida i nceteaz secreia iar submaxilara i sublinguala secret foarte puin. Alte variaii fiziologice sunt legate de vrsta subiectului, secreia crescnd pn la 20 de ani, apoi scade pn la 40 de ani, dup care se menine constant. n perioadele digestive secreia salivar crete, iar n perioadele interdigestive este redus, alimentele n funcie de specificul lor, modificnd-o att calitativ ct i cantitativ. De asemenea, n strile de deshidratare secreia salivar scade pn la 100ml/24 ore. Factorii nervoi - agitaie, team, stress, hipnoz, depresii exercit influene variabile asupra secreiei salivare, n timp ce iritaii locale i gastroduodenale, foamea, greaa, o stimuleaz. Proprietile salivei mixte Saliva este un lichid incolor, inodor, transparent, filant, spumos sau opalescent. Prin pstrarea ntr-un recipient, inferior se distinge n timp un sediment alburiu format din celulele epiteliale descuamate din canalele excretorii i cavitatea bucal; leucocite reprezentate mai ales prin polimorfonucleare i bacterii. Leucocitele ptrund n saliv prin peretele capilarelor gingivale, la nivelul anului gingivodentar, n cadrul procesului de diapedez. Ele lipsesc din saliva copiilor nainte de apariia dentiiei i de asemenea din saliva persoanelor edentate. Dintre bacteriile prezente n saliv, 70% sunt streptococi. La indivizii care prezint carii active, crete numrul lactobacililor, fr a depi 1% din totalul populaiei de bacterii. Saliva este un lichid hipoton, avnd presiunea osmotic inferioar plasmei. La un debit secretor reprezentnd 5% din debitul maxim, osmolaritatea salivei este de aproximativ 50-100 mOsm. Ea crete paralel cu debitul salivar i are tendina s devin izoton ntr-o secreie maximal. Punctul crioscopic al salivei variaz ntre -0,2 i -0,4C, tinznd s se apropie de cei al plasmei (0,56) n condiiile secreiei maximale. Densitatea salivei variaz cu glanda secretoare, condiiile n care are loc secreia, concentraia substanelor anorganice i organice, prezentnd valori cuprinse ntre 1002-1012. Vscozitatea salivei este de 1,1-1,32 n timp ce plasma prezint valori ntre 1,9 i 2,3. Reacia salivei mixte la debite secretorii mici este cuprins ntre 5,7 i 6,2. Saliva parotidian are un pH de 5,45-6,06, saliva submaxilar are un pH de 8,39. n secreia stimulat, pH-ul crete spre 8, prin creterea concentraiei de bicarbonat. La copil secreia salivar este ceva mai alcalin comparativ cu cea a adultului. 15
Reacia salivar este paralel cu reacia sngelui; pH-ul salivar crete n hiperpnee, n general n alcaloze, i scade n acidoze. Saliva se opune modificrilor brutale de pH la introducerea de substane acide sau alcaline n cavitatea bucal, prin sistemele tampon coninute: bicarbonai, fosfai, proteine, mucin, sistemul bicarbonailor avnd o pondere de 85% din puterea de tamponare a salivei. Cltirea gurii cu soluii acide, respectiv alcaline, face ca pH-ul salivei s se modifice n sens opus, ntr-un interval de timp de aproximativ 10 minute. Meninerea n limitele normale ale pH-ului salivar este important pentru crearea condiiilor de lucru a enzimelor din componena salivei, meninerea integritii structurale locale, meninerea ionilor n soluie. Scderea valorilor pH-ului salivar sub 6,5 favorizeaz apariia cariilor prin eliberarea calciului din apatit i dizolvarea materialului organic al dinilor. Creterea valorilor pH-ului peste limita superioar favorizeaz formarea tartrului dentar i al sialoliilor prin precipitarea srurilor de calciu i depunerea acestora la nivelul ductelor glandelor salivare. Compoziia salivei Se admite c saliva este alctuit din 99,4% ap i 0,6% reziduu uscat din care 0,2% sunt substane anorganice i 0,4% sunt substane organice. Compoziia salivei variaz n limite largi de la un individ la altul n funcie de stimuli, debit salivar i contribuia fiecrei glande. Dintre substanele anorganice o pondere mai mare au ionii de sodiu, potasiu, calciu, magneziu, clor, fosfat i combinaiile acestora. Alte substane anorganice sunt prezente n concentraii reduse, ca de exemplu sulful, bromul, cuprul, iodul, fluorul, cobaltul. Sodiul, n saliva "de repaus" reprezint 8,7-23,9 mEq/l. El provine din plasm, fiind secretat activ la nivelul acinului i resorbit parial la nivelul ductului striat. Concentraia sa prezint variaii n funcie de tipul de gland implicat n secreie. Saliva parotidian are cea mai nalt concentraie de sodiu, aceasta variind n funcie de debit secretor (crete la debite mari), vrst, sex (concentraia sa fiind mai mare la brbai), tip de stimulare (stimularea parasimpatica i crete concentraia). Potasiul, n saliva "de repaus" reprezint 12,8-16 mEq/l fa de 4,6 mEq/l n plasm. Potasiul este secretat activ la nivel acinar i tubular, i concentraia sa prezint variaii n funcie de: tipul de gland implicat n secreie (este mai concentrat n saliva parotidian), debitul secretor (la debite salivare mari concentraia potasiului scade prin fenomenul de diluie, n condiiile unei secreii tubulare constante) i tipul de stimulare (stimularea simpatic i crete concentraia n timp ce stimularea parasimpatica o reduce). Calciul salivar se afl ntr-o concentraie de 2,3-5,5mEq/l fa de 4,3-5,2mEq/l ct este concentraia sa n plasm. El este secretat activ la nivelul acinului. Concentraia lui n saliv depinde de tipul de glanda salivar (saliva parotidian este mai srac n calciu), de debitul salivar (concentraia calciului scade paralel cu mrirea debitului, prin creterea contribuiei parotidei), i de tipul de stimulare (stimularea simpatic crete concentraia calciului n saliv). n saliv, calciul se gsete sub dou forme: nedializabil i dializabil. Cel nedializabil este legat de proteine amilaza avnd o important contribuie i glicoproteine (dup Rolla i Jonsen o glicoprotein cu GM 100.000, cu 100 resturi de acid sialic pe molecul, leag 130 de atomi de calciu, la pH=7), iar o parte exist sub form de agregate coloidale. Saliva este o soluie saturat n fosfat de calciu, ceea ce mpiedic pierderea acestuia din smalul dentar. Exist deci, un echilibru ntre calciul i fosforul din saliv pe de o parte, i calciul i fosforul din structura cristalin a smalului dentar, echilibru care depinde i de pH-ul fluidului bucal. Valori sczute ale pH-ului favorizeaz dizolvarea hidroxi-apatitei, favoriznd apariia cariilor dentare. Fosfaii prezint n saliv concentraii de dou ori mai mari dect n plasm existnd n forma dializabil i nedializabil. Creterea debitului salivar se nsoete de scderea concentraiei fosfailor n saliv. Bicarbonaii din saliv provin din activitatea metabolic a celulelor acinare i tubulare, fiind sintetizai local n prezena anhidrazei carbonice. n secreia salivar bazal au o concentraie de 5-10mEq/l fa de 24.0-27.0mEq/l n plasm. n secreia maximal, concentraia bicarbonailor atinge 50-60mEq/l de saliv. 16
Cea mai mare concentraie de bicarbonai exist n saliva glandelor sublinguale i cea mai mic n saliva parotidian. Stimularea parasimpatic i mai accentuat stimularea simpatic cresc concentraia bicarbonailor. Fluorul salivar se afl ntr-o concentraie de 0,006-0,015mEq/l la copii i 0,005mEq/l la aduli, de acelai ordin de mrime cu cel plasmatic. Concentraia sa nu este influenat de fluxul salivar. El a fost investigat n legtur cu rolul su protector mpotriva cariei dentare i pare c, n anumite concentraii este necesar meninerii integritii dinilor deja formai. Iodul provine din plasm, prin transfer activ n poriunea tubular proximal, i are o concentraie mai mare n saliv dect n plasm. La nivelul glandelor salivare, sub influena tireotropinei hipofizare, se mai secret monoiodtirozina i diiodtirozina. Thiocianatul se gsete n saliv ntr-o concentraie mai mare dect cea plasmatic, la fumtori n concentraii mai mari dect la nefumtori. Combinat cu o protein salivar el are rol antimicrobian, bacteriostatic. Substanele organice din compoziia salivei sunt substane azotate proteice i neproteice ct i substane neazotate glucide, lipide i vitamine. Cel mai bine reprezentate, proteinele salivare, se afl ntr-o concentraie de 2,2 g/l saliv mixt "de repaus" (saliva parotidian conine 2,3 +/ 1.7 g/l; saliva submaxilar 1,2 +/ 0,8 g/l iar saliva sublingual i a glandelor salivare accesorii 2,96 g/l), concentraia proteinelor crescnd paralel cu fluxul salivar. Proteinele din saliv provin din celulele acinare glandulare, din plasm i o mic parte au ca origine microorganismele cavitii bucale. Din totalul proteinelor salivare s-au izolat 7-8 fraciuni cu proprieti imunologice i electroforetice similare proteinelor serice. Mai bine reprezentate sunt albuminele (ntre 1% i 10%) i globulinele (aproximativ 2% din proteinele totale). Imunoglobulinele sunt majoritatea IgA i mai puin IgG i IgM. Majoritatea IgA i IgM provin din glandele salivare; IgG i o mic parte de IgM provin din ser. Lactoferina este o protein fixatoare de fier, care contribuie la protejarea mucoasei bucale prin extragerea fierului necesar creterii bacteriilor din mediul bucal. n saliv exist i factorii VII, !X, VIII, III plachetari, similari cu cei plasmatici, originea lor nefiind pe deplin elucidat. Amilaza salivar sau ptialina este o alfa 1-4 glicozidaz ce hidrolizeaz legturile 1-4 glicozidice din mijlocul i din extremitile lanurilor amidonului fiert sau copt. Din aciunea enzimei rezult oligozaharide (maltoza i maltotrioza), polimeri i dextrine cu un numr mai mare de molecule de glucoza. 80% din amidon este degradat la maltoz, 20% rmnnd n stadiul de dextrine (amilodextrirte, eritrodextrine i acrodextrine). n saliv au fost izolate cinci izoamilaze cu putere hidrolitic diferit. Ptialina are pH-ui optim de aciune 6,9, temperatura optim de 37 C; este activat de ionii de Ca i Cl i inactivat de metale grele Hg, Pb, Bi ct i prin fierbere. Amilaza salivar este o glicoprotein cu greutate molecular de 50.000. Reprezint 30% din totalul proteinelor salivare n secreia parotidian. n saliva submaxilar i sublingual proporia este mai mic. Lipsete n secreia glandelor salivare accesorii. Concentraia amilazei crete proporional cu fluxul salivar i este stimulat cu precdere pe cale simpatic. Unii autori admit prezena n saliv a unei maltaze i a unei lipaze salivare (prere infirmat de unele cercetri). Lizozimul este o enzim cu structur polipeptidic. El constituie 10% din proteinele salivare. Acioneaz asupra structurilor polizaharidice din capsulele i membranele unor bacterii, avnd efect bacteriolitic. Concentraia sa variaz n limite largi de la un individ la altul i de la o determinare la alta. Activitatea lizozimic este mai mare n saliva submaxilar faa de cea parotidian. Bactericidina cu efect bactericid asupra lactobacilului a fost identificat n saliva glandelor submaxilare i a glandelor parotide. Glandele salivare produc i activatori ai kalicreinogenului care transform kalicreinogenul n kalicrein iar acesta, acionnd pe kininogen, elibereaz kininele active: kalidina i bradikinina cu efecte locale vasodilatatoare importante pentru secreia salivar. Majoritatea cercetrilor efectuate pe animale au evideniat prezena kalicreinei sub form de granule secretorii mici localizate apical n celulele ductale. 17
Mucinele sunt glicoproteine ce reprezint aproximativ 35% din totalul proteinelor salivei parotidiene i un procent mai mare n saliva submaxilar i sublingual. Mucinele confer salivei vscozitate i au rol protector n aprarea i lubrifierea mucoasei i dinilor. Funcioneaz i ca sistem tampon contribuind la meninerea pH-ului salivar. Din glandele submandibulare de oarece a fost extras factorul de cretere ai nervilor, pus n eviden i n alte esuturi (lanul ganglionar simpatic, plasm, rinichi, saliv i unele tumori). Acest factor intervine n dezvoltarea sistemului nervos, stimuleaz sistemul nervos vegetativ, crete concentraia noradrenalinei n terminaiile nervoase simpatice din miocard, glande submaxilare, stimuleaz dezvoltarea ganglionilor limfatici. Prin administrarea extractului purificat de gland submaxilar la oarecii nou-nscui s-a constatat creterea numrului de neuroni simpatici cu mrirea dimensiunilor celulare. Factorul de cretere epidermic a fost izolat n extractul de glande submaxilare la oarece. Materialul activ este o protein cu proprieti antigenice a crei activitate dispare sub aciunea enzimelor proteolitice i prin nclzire prelungit n mediul acid sau bazic. Factorul are un efect de stimulare asupra epidermei i asupra erupiei dinilor. Extractul injectat la oarecii nou-nscui produce stimularea erupiei dinilor, stimularea keratinizrii epidermei i ngroarea sa, creterea diametrului cozii, precum i modificri la nivelul epiteliilor mucoaselor cavitii bucale, esofagului i stomacului. Parotina este o substan cu rol de hormon i a fost izolat din glandele parotide, glandele submandibulare, saliv i urin. Se presupune c este secretat de celulele acinare, reabsorbit la nivelul duetelor striate, inut n spatiile intercelulare de unde prin intermediul limfei ajunge n circulaia general. Acioneaz asupra esuturilor mezenchimatoase, n special asupra esuturilor dure i esutului conjunctiv, favorizndu-le dezvoltarea. Este implicat n fixarea calciului n oase i dini, are aciune hipocalcemiant scade fraciunea nedializabil fr s modifice calciul ionic. Stimuleaz dezvoltarea cartilagiului osos i a fibrelor elastice din piele i aort, avnd efecte anabolizante n metabolismul proteic. Exist studii n care existena acestui hormon este pus sub semnul ntrebrii. Alte componente organice salivare cu rol de hormon sunt sialogastrona - produs de glandele mandibulare evideniate la obolan, cu rol inhibitor asupra secreiei gastrice, asemntoare gastronei i sialotonina - izolat n glandele salivare la pisic, cu efect vasopresor. Antigenele sistemului AB0 sunt glicoproteine prezente n saliv la 80% dintre indivizi. Aceste substane sunt secretate n saliva submaxilar i sublingual ntr-o concentraie de 15 mg/l dar nu se gsesc n saliva parotidian, n timp ce saliva glandelor accesorii labiale conine o cantitate de 6-30 de ori mai mare. Saliva poate conine i antigene ale sistemului Lewis. n saliv exist i mici cantiti de uree (n concentraie mai mare n saliva parotidian, nivelul su crescnd n uremie), acid uric (n cantiti crescute n gut) i cantiti foarte mici de aminoacizi, creatinin, amoniac. Substanele de natur glucidic din saliv sunt reprezentate pe de o parte de componentele glucidice ale glicoproteinelor i pe de alt parte de glucide libere: glucoza, galactoz, maltoz n cantiti reduse i produi ai metabolismului glucidic, acid lactic i citric. Concentraia glucozei n saliv evolueaz n paralel concentraiei sale n snge. La o valoare normal a glicemiei, glucoza n saliv este n concentraie de 0,5 mg/100ml saliv. Lipidele din saliva mixt sunt reprezentate prin digliceride, trigliceride, colesterol liber i esterificat, fosfolipide i mici cantiti de acizi grai. Compoziia salivei prezint o serie de variaii legate de modificarea fluxului salivar, contribuia fiecrui tip de gland la alctuirea salivei mixte, natura stimulrii. S-au pus n eviden variaii n funcie de ritmul circadian al concentraiilor de proteine, amilaz, sodiu, potasiu, clor, calciu, tiocianat. Nivelul de proteine, mai ridicat post prandial este corelat cu perioadele alimentare, concentraiile de Na i Cl sunt mai ridicate n primele ore ale dimineii, cele de K n cursul dup-amiezilor, iar Ca i P cresc n cursul nopii. Rolurile salivei 18
Saliva are un rol n digestie, n meninerea troficitii normale a esuturilor din cavitatea bucal, n meninerea structurii i stabilitii dinilor n alveole, dar i roluri de ordin mai general n cadrul meninerii echilibrului termic i hidroelectrolitic al organismului. n digestie, saliva se implic prin mbibarea progresiv a alimentelor n timpul masticaiei, urmat de formarea bolului alimentar. Prin aciunea de lubrifiere a mucoasei bucale, sunt favorizate masticaia, alunecarea bolului alimentar i fonaia. Prezena amilazei salivare face ca hidroliza amidonului fiert sau copt s nceap din cavitatea bucal, continund i n stomac, pn cnd bolul alimentar se mbib cu suc gastric acid, aproximativ n jumtate de or. La pH=4,5 aciunea amilazei salivare este blocat. Amilaza salivar degradeaz prin hidroliza, alimentele de natur glucidic rmase pe dini. Aciunea de protejare a esuturilor cavitii bucale se realizeaz prin: - umectarea mucoasei i tapetarea ei cu un strat fin de mucin necesar pe lng meninerea troficitii i la realizarea funciilor sale n digestie i fonaie. Aciunea sa antimicrobian se realizeaz pe de o parte prin ndeprtarea mediului favorabil creterii bacteriilor - resturile alimentare i pe de alt parte prin substanele cu aciune antimicrobian ce fac parte din compoziia salivei; lizozim, bactericidin, lactoferin, complexul globulin-tiocianat, imunoglobulinele, n special imunoglobulina A secretoare. Prin sistemele tampon coninute saliva se opune variaiilor mari de pH, meninnd saturaia sa n fosfat de calciu i mpiedicnd solubilizarea apatitei din smal. Datorit hipotoniei sale, ntre saliv i plasma sanguin rezult o deplasare de ap cu meninerea unei hidratri normale a tramei proteice interprismatice. Aceasta i ncrcarea sa electric negativ favorizat de pH-ul existent, contribuie la meninerea structurii smalului i dentinei. Secreia salivar deine un rol deosebit de important n economia general, intervenind n meninerea echilibrului hidroelectrolitic. n strile de deshidratare, reducerea coninutului de ap din organism duce la diminuarea secreiei salivare sub aciunea hormonului antidiuretic, proces urmat de uscarea mucoasei bucale i apariia senzaiei de sete. Saliva este implicat n declanarea senzaiei gustative prin solubilizarea substanelor sapide ajunse n cavitatea bucal, pe care le pune astfel n contact cu receptorii gustativi din mugurii gustativi localizai n mucoasa lingual. n saliv pot fi detectate produse toxice rezultate din reaciile metabolice: (uree, acid uric, sulfocianat); substane ajunse accidental n organism (mercur, plumb, bismut); medicamente (antibiotice, morfin); alcool etilic dar i virusuri: virusul turbrii, al poliomielitei epidemice, devenind astfel implicat n transmiterea acestor boli. MECANISMUL SECREIEI SALIVARE Saliva definitiv este rezultatul a dou procese importante, unul n care sunt implicai acinii i are ca rezultat producerea unui lichid izoton cu compoziie asemntoare unui ultrafiltrat de plasm i altul, la nivelul duetelor salivare, reprezentat de secreia i reabsorbia tubular, avnd ca rezultat saliva final hipoton. Mecanismele intime prin care fiecare substan este secretat i transferat n saliv, prezint nc necunoscute. Se poate afirma ns c exist dou laturi fundamentale ale secreiei salivare, secreia de ap i electrolii i secreia de enzime sau proteine. Numeroase studii au demonstrat c saliva primar este rezultatul unui proces de secreie activ la nivel acinar, n favoarea cruia se aduc o serie de argumente: - existena unei relaii liniare ntre consumul de oxigen, consumul de glucoza i rata secreiei de saliv; - consumuri de glucoza i oxigen mai ridicate la nivelul acinilor glandelor salivare comparativ cu metabolismul bazai (5-8 Kcal/or fa de 1 Kcal/or/Kg corp), - modificarea, n procesul secretar, a potenialului transmembranar i refacerea lui n repaus; - existena unui flux secretor acinar mpotriva presiunii superioare din ductele salivare (72 mmHg fa de 40 mmHg) la nivelul glandei parotide. 19
Apa i electroliii sunt transferate activ din plasm n lumenul acinar. Este admis existena pompelor ionice care asigur transferul ionilor n ambele sensuri att la nivelul celulelor acinare ct i la nivelul celulelor tubulare. Saliva primar, recoltat prin micropuncie n regiunea acin-duct intercalat, are presiunea osmotic i concentraia n Na+, K+ i Cl-, foarte asemntoare cu a plasmei. Rezultatele studiilor efectuate, arat faptul c la baza mecanismului de secreie al salivei se afl un mecanism de transport activ de Na+, n care sunt implicate pompa de Na+-K+, cu rolul de a menine concentraia intracelular a Na+-ului redus, a K+-ului crescut i o pomp responsabil de transportul de lichid izotonic, neutr din punct de vedere electric, care transport activ Na +, i pasiv Cl-, cuplat electrostatic cu Na+. Se presupune c pompa responsabil pentru transportul de lichid ar fi situat la nivelul membranelor celulare laterale, iar pompa Na+ - K+, att la nivelul membranei luminale ct i contraluminale. Aceasta din urm nu deplaseaz apa, dar rolul su se manifest n timpul stimulrii nervilor secretorii, cnd o cantitate important de K+ prsete celulele i este nlocuit de o cantitate echivalent de Na+. Este posibil ca influxul de Na+ s fie nsoit sau determinat de un influx de Ca2+ i o eliberare a Ca2+ din microsomii intracelulari (Peterson 1972). Calciul ar putea avea rol n cuplarea excitaiei cu secreia, n mecanismul prin care modificrile la nivelul membranei celulei acinare determin transportul de fluid la nivelul membranelor celulare laterale. O alt teorie a secreiei la nivelul acinilor glandelor salivare susine desfurarea procesului n urmtoarele etape: - stimularea nervoas - produce pe poriunea bazal a membranei celulare un efect specific i induce transportul activ al ionilor de Cl- n interiorul celulei; - creterea electronegativitii n interiorul celulei determin ptrunderea n celul a ionilor pozitivi; - excesul de ioni n interiorul celulei crete presiunea osmotic, ceea ce va aduce ap intracelular i va produce mici rupturi n marginea secretorie a ei, prin care vor trece sub presiune; ap, electroliii i materialul organic ce vor ajunge n lumenul glandei. Printre argumentele ce susin aceast teorie se afl prezena terminaiilor nervoase la nivelul polului bazal al celulelor glandulare, studiile cu microelectrozi ce arat valori ale potenialului electric al membranei polului bazal ntre 30 i 40 mV, negativ n inferior i pozitiv n exterior i care dup stimulare parasimpatic devine cu 10-20 mV mai negativ dect normal. Creterea polarizrii se produce la aproximativ o secund dup sosirea semnalului nervos, indicnd c este produs de o deplasare de ioni negativi (probabil de Cl-) prin membran, spre interiorul celulei. n timp ce Lundberg A. i colaboratorii susin c ptrunderea ionului de Cl- n celul reprezint momentul esenial n cuplarea excitaiei cu secreia, motorul secreiei de saliv, cercetri ulterioare (Petersen, Poulsen i alii) evideniaz c la baza mecanismului secretor se afl transportul de Na+ n celul, nsoit de influxul de Ca2+ extracelular i de eliberarea Ca2+ din microsomii intracelulari. n urma stimulrii vegetative, membrana celular este depolarizat de la -70mV n repaus, la 10-20mV n stare activat prin influx de Na+, concomitent influx de Ca2+ i eliberare de Ca2+ din structurile intracelulare. n 1980 Peterson arat c stimularea receptorilor colinergici, adrenergici i peptidergici determin n celulele acinare ale glandelor salivare modificri de potenial datorate creterii permeabilitii membranei pentru Na+ i K+ mediate de o cretere a concentraiei calciului n citoplasm. Dup creterea permeabilitii membranei pentru Na+ i K+, survine o hiperpolizare neasociat cu o schimbare a conductanei. Hiperpolizarea secundar se poate explica prin intrarea n joc a unei pompe electrogenice de Na+-K+, stimulat de concentraia mare intracelular de Na+. Secreia substanelor organice la nivelul celulelor acinare glandulare se caracterizeaz prin urmtoarele principii de baz deduse n urma experimentelor efectuate. Materialul necesar formrii produilor de secreie difuzeaz sau este transportat activ din capilare n poriunea bazal a celulelor glandulare, unde se afl i numeroase mitocondrii implicate n producerea de ATP. Energia obinut din ATP este folosit n sinteza de substane organice care are loc n reticulul endoplasmatic i aparatul Golgi. 20
Materialul secretat este transportat prin tubulii reticulului endoplasmatic i n aproximativ 20 de minute ajunge n veziculele complexului Golgi, situate n apropierea captului secretar al celulelor. La nivelul complexului Golgi materialul este modificat, concentrat i apoi eliberat n citoplasm sub form de vezicule secretorii depozitate la polul apical al celulelor glandulare. Aceste vezicule rmn depozitate pn cnd semnale de control umorale sau nervoase determin desfacerea lor i eliberarea coninutului la suprafaa celulei. Aceste semnale cresc permeabilitatea membranei celulare pentru calciu. Ptrunderea calciului n celul determin fuzionarea veziculelor cu membrana celular, iar ruperea lor ulterioar duce la eliberarea coninutului lor n exterior. Reabsorbia i secreia tubular Compoziia salivei finale este expresia unor fenomene complexe ce se petrec de-a lungul sistemului canalicular i care sunt parial cunoscute. Saliva primar, de-a lungul sistemului canalicular devine intens hipoton, ca urmare a unei scderi importante a concentraiei de Na+, nsoit de o cretere mai puin important a concentraiei de K+ i HCO3n timp ce cantitatea de ap nu sufer modificri semnificative. La nivelul tubilor se produce o reabsorbie activ de Na+, secreie activ de HCO3- i K+ n lumen i deplasare pasiv a ionilor de Cl- datorat gradientelor electrostatice create. Bicarbonatul total rezult din secreia activ la nivelul acinilor i la nivelul celulelor ductale striate unde se sintetizeaz n prezena anhidrazei carbonice. n condiiile secreiei unei cantiti foarte reduse de saliv, din timpul repausului secretor, mecanismele active de transport tubular ar putea fi nefuncionale sau activate la un nivel foarte sczut determinnd o secreie cu o compoziie foarte asemntoare plasmei. Prin stimularea nervilor secretori crete nivelul de activare al sistemelor transportoare pn la o limit numit transport maximal Tm - dependent de intensitatea stimulrii nervoase. Dac nivelul de stimulare are intensiti foarte mari i acinii glandulari furnizeaz cantiti mari de saliv primar, este afectat n primul rnd mecanismul de transport al Na+-ului i ca urmare, se produce creterea concentraiei sodiului n saliva final. REGLAREA SECREIEI SALIVARE Reglarea secreiei salivare se face n principal pe cale nervoas, prin reflexe necondiionate i condiionate, dar i sub influena factorilor umorali, cu rol corector i integrator n funcionarea general a organismului. Reflexele necondiionate au ca suport arcul reflex format din receptori, ci aferente, centrii nervoi i ci eferente. Receptorii sunt localizai n principal n cavitatea bucal i sunt reprezentai de receptori gustativi, mecanoreceptori, termoreceptori i algoreceptori. n afara acestora sunt stimulai i mecanoreceptorii din articulaia temporo-mandibular, proprioreceptorii din muchii masticatori, receptorii din mucoasa esofagului, stomacului i chiar din afara tubului digestiv. Fibrele aferente ce preiau impulsurile de la receptorii gustativi localizai n cele 2/3 anterioare ale limbii intr n alctuirea nervului coarda timpanului, fac apoi cale comun cu lingualul pentru ca n final, prin nervul intermediar Wrisberg s ajung la centru salivator superior din formaiunea reticulat pontin. De la nivelul receptorilor gustativi ai 1/3 posterioare a limbii, impulsurile sunt preluate de fibre aferente ce intr n alctuirea glosofaringianului i ajung n centrul salivator inferior, localizat n substana reticulat bulbar. Aferente de la restul receptorilor implicai n reflexe salivare sunt cuprinse n ramurile senzitive ale nervului trigemen i conduc impulsuri mai nti la nucleul senzitiv al trigemenului i apoi la nucleii salivari. Tot pe cale reflex, secreia salivar poate fi declanat prin stimularea unor receptori aflai n afara cavitii bucale; astfel, reflexul esofago-salivar de hipersalivaie, este declanat pe cale vagal, ca urmare a distensiei i iritaiei locale a esofagului n cazul existenei unor leziuni tumorale; reflexul gastro-salivar de 21
hipersalivaie este iniiat tot pe cale vagal, n cazul unor iritaii locale gastro-duodenale sau de nsi prezena alimentelor n stomac. Centrii reflexului salivar (de salivaie) sunt reprezentai printr-un grup de neuroni localizai ntre nucleii facialului i nucleul ambiguu n regiunea dorsolateral a substanei reticulare din poriunea caudal a punii i jumtatea rostral a bulbului. n apropierea nucleului ambiguu, n poriunea rostral a bulbului se afl nucleul salivator inferior. n poriunea caudal a punii, n vecintatea nucleului senzitiv al facialului se afl nucleul salivator superior. Centrii salivatori sunt conectai cu centrii superiori, hipotalamici i corticali. Stimularea electric a hipotalamusului anterior activeaz mecanismele de termoliz asociate cu hipersalivaie. Stimularea hipotalamusului posterior, n stri de stress emoional puternic, se nsoete hiposalivaie. Exist de asemeni, conexiuni ntre centrii salivatori i centrii hipotalamici ai foamei i saietii implicate n comportamentul alimentar. Hipersalivaia s-a obinut i prin stimularea zonelor corticale de proiecie a gustului, mirosului sau percepiei tactile din regiunea cavitii bucale. Din nucleul salivator superior pornesc fibre eferente parasimpatice preganglionare. Acestea intr n alctuirea nervului intermediar Wrisberg, strbat ganglionul geniculat fr sinaps, ajung n nervul coarda timpanului care se va uni cu nervul lingual pentru ca, n planeul bucal, fibrele destinate glandelor sa prseasc nervul lingual i s fac sinaps cu neuronii postganglionari din ganglionul Lagley i mai muli ganglioni din hilul glandelor submaxilare, de unde se distribuie acestora. Fibre postganglionare scurte se distribuie glandelor sublinguale. Din nucleul salivator inferior pornesc fibre parasimpatice preganglionare ce intr n alctuirea nervului glosofaringian, strbat ganglionul Andersch fr sinaps, intr apoi pe rnd, n alctuirea nervului Jacobson, nervului micul pietros superficial iar n ganglionul otic fac sinaps cu neuronii postganglionari parasimpatici. Fibrele postganglionare scurte ajung pe calea nervului auriculotemporal, n glanda parotid. Fibrele eferente simpatice i au originea n neuronii simpatici din coarnele laterale medulare ale regiunii toracale T1 i T2. Fibrele simpatice preganglionare prsesc mduva pe calea rdcinilor anterioare ale nervilor spinali i prin lanul simpatic paravertebral ajung n ganglionii cervicali superiori unde fac sinaps cu neuronii postganglionari. Fibrele postganglionare lungi ajung la glandele salivare pe traiectul vaselor. Reflexele condiionate salivare iau natere prin asocierea n timp ntre stimulul necondiionat alimentul - i stimulii condiionai (olfactivi, vizuali, mintali). De la nivelul receptorilor impulsurile ajung pe ci aferente, diferite cu natura stimulului condiional, n arii de proiecie specifice conectate cu arii corticale i subcorticale ce influeneaz secreia salivar. Eferenele conduc impulsuri secretorii, elaborate n structuri nervoase superioare i, prin centrii salivari medulo-bulbo-protuberaniali pn la glandele salivare. Se realizeaz, pe aceast cale, integrarea secreiei salivare n reaciile de adaptare ale organismului. Primul care a dovedit experimental existena acestui tip de reflex a fost I.P.Pavlov (1936) care a asociat, n mod repetat, un stimul indiferent cu alimentaia, conferindu-i calitatea de stimul condiionat; ulterior este suficient aplicarea numai a excitantului condiionat pentru a se declana reflex salivaia. Totui, dac asocierea celor dou tipuri de excitani nu este rentrit, reflexul salivator se stinge. Au fost descrise trei faze ale secreiei salivare, controlate reflex n mod diferit: faza cefalic (secreia salivar este stimulat de vederea, mirosul, sau gndul la un aliment, fiind declanat naintea introducerii acestuia n cavitatea bucal); faza bucal (secreia salivar este declanat de contactul alimentelor cu mucoasa oral i stimularea receptorilor acesteia); faza gastro-intestinal (declanat reflex de prezena alimentelor n stomac). Efectele stimulrii simpatice si parasimpatice asupra secreiei salivare Glandele salivare primesc att fibre simpatice ct i parasimpatice. Acestea inerveaz diferitele tipuri de celule secretoare dar i vasele de snge i celulele mioepiteliale contractile, inducnd interaciunile ntre procesele secretorii, vasomotorii. Activitatea secretorie a glandelor salivare este controlat mai ales prin intermediul fibrelor parasimpatice. Impulsurile parasimpatice se transmit celulelor efectoare prin intermediul 22
acetilcolinei ce acioneaz pe receptorii M-colinergici din membranele celulelor efectoare i produc eflux de K+, influx de Na+, hiperpolarizarea membranei celulare, acumularea de Ca2+ n celul. Rezultatul va fi producerea unei salive abundente, fluid, bogat n Na+, Cl-, HCO3- srac n K+ Ca2+, proteine. Stimularea parasimpatic intensific, dar mai puin dect stimularea simpatic, secreia de enzime i de mucus. Simultan, se produc vasodilataie i creterea consumului de oxigen, ambele neinfluenate de administrarea de atropin, ceea ce indic implicarea altor mecanisme - eliberarea local de kalicrein cu producere de kinine active vasodilatatoare. Stimularea fibrelor simpatice prin aciunea noradrenalinei i adrenalinei care acionnd pentru receptorii a-adrenergici produc creterea secreiei de K+ i ap a celulelor acinare, iar stimularea receptorilor (3-adrenergici produce eliberarea enzimelor din granulele de zimogen, ale celulelor acinare. Stimularea simpaticului produce o saliv n cantitate redus, vscoas, bogat n proteine, cu o concentraie crescut a ionilor de K+, Ca2+,HCO3- i mai redus de Na+ i Cl-. Simpaticul produce i contracia celulelor mioepiteliale i vasoconstricie, urmat secundar de vasodilataie produs de kininele vasodilatatoare i de acumularea local de metabolii. La nivel tubular simpaticul i parasimpaticul au efecte asemntoare, modificnd transportul prin peretele tubular. Influene hormonale asupra secreiei sitroficitii glandelor salivare Secreia i dezvoltarea glandelor salivare este influenat i de hormonii unor glande endocrine, cum ar fi: hormonii tiroidieni (au aciune trofic nespecific asupra glandelor, iar tiroidectomia este urmat de atrofia glandelor salivare); hormonii adenohipofizei, STH, TSH i ACTH au de asemeni aciune trofic nespecific (hipofizectomia la obolan produce atrofia glandei parotide); aldosteronul secretat de corticosuprarenal stimuleaz reabsorbia de Na+ i eliminarea de K+ la nivelul ductelor glandulare (aciune asemntoare celei de la nivel renal); ADH-ul hipotalamic reduce pierderea de ap prin saliv (similar aciunii de la nivel renal), explicndu-se astfel hiposalivaia din strile de deshidratare. Alte substane ce intervin n reglarea secreiei salivare sunt angiotensina, bradikinina, atropin, ergotamina, cu efect inhibitor asupra secreiei salivare; adrenalina, acetilcolina, pilocarpina, histamina cu efect stimulator asupra secreiei salivare. Factorii umorali intervin n secreia salivar cu rol corector, integrator n unele funcii homeostazice, sau trofic asupra glandelor salivare. Rspunsurile secretorii difereniale i specificitatea secreiei salivare n raport cu stimulul aplicat dovedesc complexitatea actului de control i integrare a secreiei salivare. Exist o specificitate a secreiei salivare n raport cu stimulii prezeni n cavitatea bucal, astfel: alimente uscate (pesmet, pine uscat), determin producerea unei salive apoase, abundente, srac n compui organici, substanele acide declaneaz o secreie salivar bogat, cu mult mucus; carnea produce o saliv cu vscozitate crescut etc.
IV. SENSIBILITATEA GUSTATIV Gustul, mpreun cu mirosul, sunt implicate n identificarea alimentelor nocive pentru organism i permite unui individ s selecteze hrana conform dorinelor i poate necesitilor metabolice n anumite circumstane. El reprezint o senzaie complex, iniiat prin stimularea receptorilor gustativi dar i tactili, termici, olfactivi de la nivelul mucoasei linguale i buco-faringiene, urmat de transmiterea impulsurilor pe ci de conducere aferente la centrii gustativi aflai n interrelaie cu ali centri, n vederea hrnirii. Principalii receptori ai gustului, mugurii gustativi, sunt situai la nivelul papilelor fungiforme, circumvalate i foliate ale limbii, dar i la nivelul membranei mucoase a faringelui i n mod particular, ia 23
nivelul epiglotei. Adulii au un numr de aproximativ 10.000 muguri gustativi. Peste 45 de ani, numrul lor se reduce, scznd i acurateea senzaiei gustative. Cele mai numeroase i mai mici, papilele filiforme sunt distribuite pe toat faa dorsal a limbii din poriunea anterioar i nu conin muguri gustativi. Papilele fungiforme, n form de ciuperci, se afl tot pe faa dorsal a limbii, printre papilele filiforme. Ele sunt alctuite dintr-un miez de esut conjunctiv i la suprafa au un epiteliu stratificat scuamos. Pe faa dorsal se afl un numr moderat de muguri gustativi. n afeciunea numit disautonomia familial, lipsesc papilele fungiforme, mugurii gustativi i gustul. Papilele circumvalate n numr de 7-9 sunt dispuse n "V", paralel cu sulcus terminalis. Aceste papile sunt mari i rotunde. Ele au miezul format din esut conjunctiv, acoperit de un epiteliu stratificat scuamos. Sunt nconjurate de un an adnc al crui epiteliu conine muguri gustativi. Pe marginea limbii sunt localizate papilele foliate care conin un numr moderat de muguri gustativi. n seciune histologic, mugurele gustativ apare ca un con oval, palid colorat, cu axul lung strbtnd grosimea epiteliului. Vrful mugurelui gustativ atinge suprafaa epiteliului. Accesul n cavitatea bucal se realizeaz printr-o mic deschidere n epiteliu numit por gustativ. Structura mugurelui gustativ este reprezentat prin trei tipuri de celule. Dou dintre ele, celulele de susinere i celulele senzoriale sau neuroepiteliale, sunt alungite i sunt cuprinse ntre lamina bazal i porul gustativ. Extremitatea celular din dreptul porului gustativ se ngusteaz i fiecare celul trimite prin porul gustativ microvili, pe suprafaa receptoare a gustului, ce ajung n contact direct cu saliva i substanele sapide. Ei sunt nconjurai de o substan amorf, dens, presupus a fi produsul de secreie al celulelor de susinere - acelai material poate fi observat i n interiorul acestor celule. Ai treilea tip, celulele bazale, se afl la periferia mugurelui gustativ, lng lamina bazal. Se presupune c acestea dau natere celorlalte dou tipuri celulare a cror medie de via este de aproximativ zece zile. La baza mugurelul, sub membrana bazal se realizeaz o bogat reea de fibre nervoase senzitive ce aparin perechilor VII i IX de nervi cranieni, formnd plexul gemal. Ramuri ale acestui plex nconjoar mugurele gustativ, formnd plexul perigemal i ntre muguri - plexul intergemal, din care se desprind filete nervoase terminale care ptrund n mugure i formeaz plexul intragemal. Aceste filete sunt destinate celulelor gustative, cu care realizeaz sinapse speciale neuroepiteliale. n interiorul mugurelui gustativ ptrund i fibre amielinice cu origine simpatic. Distrugerea fibrelor senzitive duce la degenerarea mugurilor sau dispariia lor, n timp ce regenerarea nervului determin reapariia mugurilor ceea ce dovedete influena trofic a terminaiilor nervoase asupra celulelor gustative. Fiecare fibr senzitiv colecteaz informaii de la mai multe celule gustative i realizeaz o convergen a stimulilor. Mugurii gustativi sunt localizai n arii speciale n funcie de senzaia gustativ pe care o deservesc. Pentru gustul dulce i srat, mugurii gustativi sunt localizai spre vrful limbii, gustul acru este deservit de mugurii gustativi localizai pe prile laterale, iar gustul amar de cei localizai n partea posterioar a limbii i pe palatul moale. Studiile efectuate pe mugurii gustativi unici pe care se exercit aciunea fiecruia dintre cei patru stimuli gustativi primari, arat c cei mai muli rspund stimulrii cu doi, trei i chiar patru stimuli, precum i a altor tipuri de stimuli gustativi ce se ncadreaz n primele patru categorii. Generarea senzaiei gustative se produce ca urmare a aciunii exercitate a substanelor sapide dizolvate n saliv. Ele produc depolarizarea membranei microvililor celulelor gustative. Scderea potenialului este aproximativ proporional cu logaritmul concentraiei substanei stimulante. Aceast schimbare de potenial la nivelul celulelor gustative reprezint potenialul de receptor. Mecanismul prin care acesta este iniiat, const probabil n crearea unor legturi ntre substana absorbit i proteinele receptoare ale membranei microvililor, cu deschiderea unor canale ionice care permit ionilor de Na+ s ptrund i s depolarizeze celula. Dac potenialul de receptor atinge un anumit prag, se transform n potenial de aciune. Pe msur ce substana sapid este splat de saliv, senzaia gustativ dispare prin ndeprtarea stimulului.
24
Exist ipoteze care admit formarea unor compui chimici ntre stimul i receptorii specifici de natur proteolipidic de pe membrana receptoare ce modific permeabilitatea membranei i genereaz potenialului de receptor care, la o valoare prag, iniiaz potenialul de aciune. Alt ipotez admite existena pe suprafaa microvililor a unei pelicule n care ionii substanelor sapide produc modificarea configuraiei spaiale a distribuiei sarcinilor electrice i generarea potenialului de receptor. O prim aplicare a stimulului gustativ se nsoete de un vrf al ratei de descrcare a impulsurilor n fibrele nervoase pentru ca dup 2-3 secunde s se ajung la un nivel inferior, constant, indicnd un fenomen de adaptare. Adaptarea devine mai rapid i mai complet dac excitantul are intensitatea mai mic i diminua cu sporirea intensitii excitantului. S-a constatat experimental o adaptare mai rapid pentru substanele srate i dulci i o mai lent adaptare pentru cele amare i acre. Pentru o senzaie gustativ, alimentul trebuie s fie solubil n saliv i s aib o concentraie minim, de prag. Pragurile au valori diferite n funcie de substan, de topografie i de numrul mugurilor gustativi excitai, fr a exista ns o separare net. Cel mai sczut prag s-a constatat pentru stimulii care conduc la senzaia de gust amar. Pragul gustativ este influenat de o serie de factori cum ar fi: vrsta (copii au o sensibilitate mai mare), suprafaa de stimulare, compoziia alimentelor, temperatura (maximul de sensibilitate este prezent pentru o temperatur a soluiilor de 30-40C), foamea (induce scderea pragului pentru dulce i srat i creterea pragului pentru amar i acru), lumina (sensibilitatea gustativ scade la ntuneric), existena variaiilor circadiene (creterea sensibilitii gustative seara). Pragul gustativ poate fi influenat i de unele condiii patologice cum sunt: insuficiena cortico-suprarenal, pseudoparatiroidismul etc., senzaia gustativ avnd ns i o important component individual ctigat prin antrenament. Pentru a crea senzaia gustativ, impulsurile sunt culese din 2/3 anterioare ale limbii prin nervul coarda timpanului ram din facial, iar prin nervul glosofaringian din 1/3 posterioar a limbii. Primul neuron al cii gustative se afl n ganglionul geniculat (pentru coarda timpanului) i n ganglionul Andersch i Ehrenritter (pentru glosofaringian). De la acest nivel impulsurile sunt transmise la nucleul tractului solitar din bulb unde se afl al doilea neuron al cii gustative. Apoi, pe calea nervului vag ajung i impulsurile gustative de la baza limbii, faringe, palatul moale, epiglot i laringe. Dup ce prsesc nucleul tractului solitar, fibrele cii gustative trec de partea opus i urc pe calea lemnisculului median (banda Reil) pn la al treilea neuron, situat n nucleul dorsal postero-median din talamus. Proiecia cortical a sensibilitii gustative se realizeaz n zona inferioar a circumvoluiei parietale ascendente (aria 43), n apropierea centrilor masticatori, ai deglutiiei i centrilor motori ai limbii. Un numr mare de impulsuri din tractusul solitar ajunge direct la nucleii salivatori pontini i bulbari de unde sunt transmise impulsuri secretoare ctre glandele parotide (pe calea nervului gloso-faringian, de la nucleul salivator inferior din bulb) i ctre glandele submandibulare i sublinguale (pe calea nervului facial, de la nucleul salivator superior din punte). Tot la nivelul tractului solitar pornesc ramuri colaterale spre nucleul reticulat al formaiunii reticulate de unde, pe calea tractului reticulo-bulbar se transmit impulsuri la glandele salivare, muchii pieloi ai feei, la muchii masticatori implicai n deglutiie i la muchii limbii, n scopul coordonrii micrilor asociate senzaiilor gustative. V. SENSIBILITATEA EXTEROCEPTIV I PROPRIOCEPTIV LA NIVELUL CAVITII BUCALE Sensibilitatea tactil Structurile sistemului orofacial sunt dotate cu o mare sensibilitate la diveri stimuli din mediul extern, inclusiv la stimulii dureroi. Sensibilitatea tactil cu cele trei variante - de atingere, de presiune i vibratorie include receptori din categoria mecanoreceptorilor rspndii n piele, n mucoasa oral i n capsula articulaiilor. Dei nu exist totdeauna o difereniere morfologic sesizabil, receptorii prezint o specializare funcional nendoielnic, o adaptare a structurilor morfologice de a rspunde predilect la un anumit tip de 25
energie. Ei prezint un prag foarte sczut pentru variaia unei anumite forme de energie i rmn insensibili la intensiti normale ale altor stimuli senzoriali. n cadrul sensibilitii tactile deosebim sensibilitatea la atingeri uoare, la presiune i vibraii, senzaii determinate de intensiti diferite ale aceluiai stimul de baz. Senzaia tactil este determinat de stimularea receptorilor specifici din tegument sau esuturi supraadiacente; senzaia de presiune este rezultatul deformrii esuturilor profunde; senzaia vibratorie este determinat de stimulri repetitive rapide. n stratul submucos al mucoasei bucale exist o reea bogat de fibre nervoase, terminaii nervoase libere i corpusculi senzitivi, receptori ai sensibilitii exteroceptive tactil - presional, termic i dureroase. Zonele cu sensibilitate tactil maxim sunt buzele, vrful limbii, gingiile i mucoasa palatului dur. O sensibilitate tactil mai redus o reprezint mucoasa obrajilor. Receptorii care deservesc aceste sensibiliti sunt mecanoreceptori. Mecanoreceptorii din mucoasa bucal sunt stimulai de presiunea i deformarea elastic a esuturilor locale. Introducerea alimentelor n gur, genereaz stimuli implicai n reglarea masticaiei, salivaiei i deglutiiei. - Corpusculii lui Meissner, formai din terminaii nervoase ncapsulate, servesc la recunoaterea exact a punctului stimulat i la recunoaterea texturii obiectelor cu care vin n contact. Sunt receptori ca adaptare rapid - emit impulsuri numai cteva milisecunde dup aplicarea stimulului, chiar dac stimularea se menine constant. Sunt foarte sensibili la micarea obiectelor uoare i la vibraii cu frecvene joase, n jur de 30 de Hz. Ei se dezvolt n jurul terminaiilor fibrelor de tip A, cu diametrul de 5-15, i viteza de transmitere de 20-90 m/sec. - Discurile lui Merkel semnalizeaz atingerea continu a obiectelor cu pielea. Dup ce au fost stimulate, induc o descrcare puternic de impulsuri n fibra senzitiv, apoi se adapteaz parial i transmit semnale mai slabe, dar continui. Se dezvolt n jurul fibrelor de tip A. - Corpusculii Ruffini pot recepiona pe lng stimulii calzi i pe cei tactili de presiune. Sunt formai din terminaii arborizate. Ei sesizeaz deformri puternice continue i prelungite. Adaptarea lor este redus. - Corpusculii Pacini sunt formai din lamele conjunctive concentrice care nconjoar terminaia nervoas amielinic a unei fibre A. Ei servesc detectrii schimbrilor foarte rapide n starea mecanic a esuturilor. Sunt stimulai de presiune i de micrile foarte rapide, chiar i de intensitate foarte slab. Sunt sensibil la vibraii cu frecvene cuprinse ntre 60-500 Hz; prezint o adaptare foarte rapid. - Receptorii tactili formai din terminaii nervoase libere, aparin fibrelor de tip A cu viteza de conducere de 5-30 m/sec, i fibrelor amielinice de tip C, cu viteza de transmitere de 0,5-2 m/sec. Att n cazul tactului ct i al presiunii, variaia intensitii stimulului tactil genereaz senzaia de presiune iar creterea intensitii acestuia genereaz senzaia de durere. Valoarea de prag a unui stimul este influenat de temperatur (crete cu temperatura) i de modalitatea de stimulare - pragul este mai mic dac stimulul prezint oscilaii sau micri. Intensitatea stimulului eficient depinde i de structura i situaia receptorilor - pentru corpusculii Meissner situai mai superficial i formai din mai puine straturi de esut conjunctiv, intensitatea stimulului ce produce deformarea este mai mic fa de aceea necesar stimulrii corpusculilor Pacini situai mai profund i cu structura mai bogat n esut conjunctiv. Un alt factor ce influeneaz valoarea intensitii stimulului eficient este densitatea receptorilor. Cu ct aceasta este mai mare, discriminarea stimulilor este mai fin. n ligamentul periodontal s-au descris fibre'cu diametrul mai mare, mielinice, care se termin sub form de buton (Lewinky i Steward, 1937); spiral strns (Rapp i colaboratorii, 1957); fusuri (Bernick, 1966) sau arborizaii (Simpson, 1966); implicate probabil n senzaia tactil i fibre cu diametrul mic, mielinice i amielinice care par a se termina n arborizaii fine i sunt implicate probabil n sensibilitatea dureroas. De asemeni, mecanoreceptorii perioclontali au fost studiai electrofiziologic la animalele de experien, prin nregistrarea potenialelor de aciune, din fibrele nervilor dentari, n urma aplicrii pe dini a unor fore cunoscute. S-au evideniat mecanoreceptori cu adaptare lent, care la aplicarea unui stimul constant produc descrcri continue cu o mic scdere a frecvenei i cu adaptare rapid care produc numai 26
una, dou poteniale, imediat dup aplicarea stimulului sau, nu rspund deloc, dac viteza de aplicare a stimulului nu este suficient de mare (Hannam, 1969). Unele uniti mecanoreceptoare cu adaptare lent prezint descrcri spontane n absena unui stimul vizibil, iar aplicarea stimulului de presiune crete frecvena lor de descrcare (Hannam, 1969). Sensibilitatea mecanoreceptorilor periodontali se modific odat cu direcia de aplicare a forei. Rspunsul receptorului scade progresiv atunci cnd direcia de aplicare a forei se modific spre 180 fa de direcia optimal (Pfaffmann, 1939). Receptorii din ligamentul periodontal sunt parte din cei implicai n masticaie. Ei particip n evocarea senzaiei tactile i de presiune de la nivelul dinilor care stau la baza simului ocluzar i al capacitii stereognostice de la acest nivel, La dinii normali, pragurile tactile sunt de 0,44 g pentru incisivi (Willkic,1969; Garton, 1968); 1,32 g pentru canini (Willkic,1969; Garton, 1968); 6-7 g pentru molari (Loewenstein i Rath Kamp 1955). Depulparea crete pragul cu 57% iar aplicarea unei coroane metalice cu 127%. Puterea de discriminare ntre obiecte cu diferite grosimi i forme aflate ntre suprafeele ocluzate este maxim cnd se aplic fore ntre 50 i 500 g fr ca mecanoreceptorii s fie singurii implicai. Anestezia local a ambelor maxilare reduce evident capacitatea de discriminare de la 60 m grosime la 150 m. Frecvena rspunsului receptorilor este proporional, n anumite limite, cu intensitatea i viteza de aplicare a stimulului. Viteza de conducere a fibrelor senzitive este cuprins ntre 25-28 m/sec. Aceste fibre sunt amestecate n nervii dentali cu fibre senzitive de diametru mare, provenite din piele. Receptorii din articulaia temporo-mandibular apar fie sub form de terminaii nervoase libere rspndite n ntreaga capsul cortical, fie sub form de receptori Ruffini, Vater-Pacini i Golgi, modificai, aflai n partea lateral a capsulei i n ligamentul lateral (Thiiander,1961; Ishibashi, 1966; Kawamuri i Majima, 1964). Pe lng rolul de nociceptori, unii dintre receptori par a fi implicai i n semnalizarea poziiei condilului. Prin injectarea de anestezice locale n articulaie s-a evideniat faptul c receptorii articulari joac un rol important n percepia poziiei mandibulare (Thilander, 1961). Senzaia de durere n articulaia temporo-rnandibular poate inhiba puternic micrile condilului. Informaiile senzitive de la nivelul articulaiei temporo-mandibulare particip la controlul micrilor de ocluzii. Studii de neurohistologie i de electrofiziologie au evideniat n articulaia temporo-mandibular patru tipuri de receptori: - tipul I de receptori, corpusculii Ruffini, se afi n straturile superficiale ale capsulei articulare. Fiecare receptor controleaz o anumit poziie aparinnd unui unghi cu deschidere de 15-30; - tipul II de receptori, corpusculii Pacini, se afl n straturile profunde ale capsulei articulare i n esutul celulo-grsos periarticular. Sunt mecanoreceptori dinamici, cu adaptare rapid, care informeaz asupra accelerrii micrii mandibulei; - tipul III de receptori, corpusculii Golgi din ligamentele articulare, sunt mecanoreceptori dinamici cu adaptare lent i prag de sensibilitate ridicat. Aceti receptori sunt indicatori de poziie i de micare n articulaie, informnd i asupra presiunilor ridicate din articulaie; - tipul IV de receptori, sunt terminaiile nervoase libere, rspndite difuz n structurile articulaiei emporo-mandibulare exceptnd sinoviala i meniscul articular. Unele fibre sunt nemielinizate, cu diametru de 2 microni sau puin mieiinizate cu diametrul de 2-5 microni i reprezint algoreceptori neadaptabili, altele sunt mietinizate, au diametrul mai mare i sunt mecanoreceptori care recepioneaz tensiunea, presiunea i micarea n articulaie. Sensibilitatea termic Receptorii senzaiilor termice, rspndii n toat mucoasa bucal au densitatea mai mare la nivelul buzelor fa de limb i mai ridicat la nivelul limbii fa de celelalte zone. Gradientele termice sunt discriminate prin receptori pentru rece, receptori pentru cald, nociceptori pentru rece i nociceptori pentru cald, ultimele dou tipuri fiind stimulate doar de valori termice duntoare organismului. 27
n mod clasic, receptorii pentru rece au fost considerai corpusculii Krausse. Studii mai recente au artat c axonii implicai n recepia termic i pierd teaca de mielin mai timpuriu dect mecanoreceptorii. Terminaiile receptoare sensibile la nclzire uoar aparin probabil fibrelor C, nemielinice, cu viteza de conducere de 0,5-1,5 m/sec. Cmpurile lor receptoare au diametrul mai mic de 1 mm. Terminaiile receptoare pentru rece sunt asociate cu fibre senzitive AS, cu viteza de conducere de 1420 m/sec. Cmpul receptor ai acestor fibre are diametrul de 1 mm. Numrul receptorilor pentru rece este de 3-4 ori mai mare dect ai receptorilor pentru cald. Sensibilitatea dureroas Durerea reprezint un sistem de alarm, o modalitate de semnalizare a aciunii unor ageni nocivi organismului, un mecanism complex, constnd ntr-o suocesiune de procese fiziologice i psihice ncepute pe locul aciunii stimulului nociv unde mesajele sunt codificate, transformate n poteniale de aciune, transmise centrilor nervoi unde vor fi prelucrate i transformate n senzaie contient. Receptorii nociceptivi sunt terminaii nervoase libere ale fibreior mielinice subiri de tip A ,cu viteze de conducere n jur de 15 m/sec i ale fibrelor amielinice de tip C, cu viteze de conducere n jur de 1 m/sec. Aceste fibre au proprietatea de a rspunde numai stimuiilor nocivi organismului, presiunii duntoare, ageni chimici - genereaz durere la un pH tisular de 6,2, durerea fiind maxim la pH = 3,2; stimulii termici care depesc 45C sau sunt foarte reci producnd leziuni la nivelul mucoasei bucale sau a esuturilor profunde. Cei mai muli dintre receptori s-au dovedit a fi polinodali, api s rspund mai multor categorii de stimuli. Ei acioneaz ca transductori biologici ,convertind o form de energie a stimulului, n impuls nervos. Un stimul algogen suficient de intens va produce creterea permeabilitii membranare pentru Na+, care ptrunde n celul n timp ce ionii de K+ ies n exterior, la natere astfel potenialul de receptor de tip electrotonic, cu posibiliti de sumaie. Pentru o intensitate mic a stimulului nociceptiv va lua natere un potenial de receptori de civa mV, care nu se propag. Un stimul dureros mai puternic va genera un potenial de aciune modulat n frecven care se propag slttor de-a lungul fibrei nervoase. Receptorii nociceptivi sunt capabili s semnalizeze intensitatea stimulului prin creterea frecvenei impulsurilor aferente n fibra senzitiv, n funcie de intensitatea stimulrii. Codificarea distribuiei spaiale a stimuiilor se datoreaz somatotopiei corticale. Momentul n care receptorul ncepe s descarce nu coincide cu momentul perceperii durerii, ceea ce sugereaz o sumaie temporal i spaial a impulsurilor nociceptive la nivelul sistemului nervos central. Receptorii nociceptivi nu se adapteaz, observndu-se n anumite condiii scderea pragului dureros. Tipuri particulare de durere La nivelui regiunii orofaciale se poate vorbi despre mai multe tipuri particulare de durere. Durerea de origine vascular rezult n urma lezrii vaselor sau esutului perivascuiar. Vasele de snge sunt prevzute cu chemoreceptori stimulai de substane productoare de durere ca bradikinina, histamina, serotonina etc. Stimularea receptorilor vasculari intrapulpari conduce la eliberarea de substane productoare de durere. ntre momentul aciunii stimulului dureros i apariia potenialului de aciune n fibrele senzitive, exist o perioad de laten. Se mai constat prezena fenomenului de postdescrcare - prelungirea descrcrii fibrelor nervoase dup ncetarea aciunii stimulului, timp necesar neutralizrii substanelor eliberate, productoare de durere. Durerea de origine salivar este consecina lezrii glandelor salivare mai ales prin formare de sialolii. Se produce distensia sistemului canalicular prin acumularea salivei datorat obstruciei prin sialolit. Mai frecvent afectat este glanda submandibular, durerea manifestndu-se n cursul alimentaiei. Durerea muscular este cel mai frecvent produs prin contracie muscular spontan. Ea are un caracter profund, surd, deseori iradiind n regiunea oro-facial. Cel mai important rol n producerea durerii l au deficitul de oxigen, modificrile valorilor de pH, acumularea de acid lactic, K+, histamina. Durerea facial este produs ca urmare a spasmului, ischiemiei, inflamaiei musculaturii maxilare, diverse perturbri la nivelul tendoanelor, ligamentelor, articulaiei temporo-mandibulare. 28
Segmentul intermediar i central al sensibilitii extero i proprioceptive de la nivelul cavitii bucale - prin ramuri trigeminale. Fibrele ce conduc sensibilitatea tactil de la nivelul mucoasei cavitii bucale i de la nivelul parodoniului sunt dendrite ale neuronilor din ganglionul lui Gasser, axonii lor formeaz rdcina pontin a trigemenului ce se termin n nucleul pontin trigeminal. Sensibilitatea termic i dureroas de la nivelul cavitii bucale, din dini i din articulaia temporomandibular este condus de ctre fibrele cu originea n ganglionul senzitiv al lui Gasser; ele formeaz rdcina descendent sau spinal a trigemenului care se termin n nucleii trigeminali din bulb i n primele segmente cervicale medulare. n nucleul interpolaris din bulb se termin fibrele termice, iar n subnucleul caudalis se termin fibrele sensibilitii dureroase. Fibrele ce conduc sensibilitatea proprioceptiv de la nivelul muchilor masticatori ct i presiunile exercitate asupra mecanoreceptorilor periodontali i articulari formeaz rdcina mezencefalc a trigemenuiui,avnd originea n neuronii senzitivi pseudounipolari din nucleul mezencefalic al trigemenului. De la nivelul neuronilor din nucleii senzitivi ai trigemenului, fibrele ascendente ale sensibilitilor li se altur i cele medulare. Pentru toate aceste ci al treilea neuron este situat n talamus de unde axonii lor se proiecteaz n scoara cerebral, lobul parietal, zonele 3, 1, 2. Prin conexiunile pe care le realizeaz cu neuronii motori din trunchiul cerebral i cu nucleii salivatori din bulb i din punte, neuronii senzitivi ai trigemenului nchid cercul reflexelor masticatorii i de deglutiie i influeneaz procesul secreiei salivare. VI. ARTICULAIA TEMPORO-MANDIBULAR Constituie articulaia mobil de legtur dintre baza craniului i mandibul, permind micarea suprafeelor ocluzale ale dinilor antagoniti situai pe cele dou arcade. Articulaia temporo-mandibular este o articulaie condilian, format din condilul mandibulei, condilul temporal i cavitatea glenoid. ntre cele dou suprafee articulare se gsete meniscul, care mparte articulaia n dou compartimente: unul superior sau temporomeniscal i unul inferior, meniscomandibular. Suprafeele meniscului sunt acoperite de o sinovial ce dubleaz faa intern a capsulei articulare. Capsula articular, ntrit de ligamente nvelete toate elementele articulare, ca un manon fibroelastic. Laxitatea ligamentelor i lipsa crestelor osoase n jurul cavitii glenoide permit micri foarte variate n articulaia temporo-mandibular. Ele sunt micri n direcia sagital, transversal sau vertical, n raport cu tipul dinamic masticator. Micarea de translaie a condilului i a meniscului, asociat cu micarea de rotaie a lui n timpul deschiderii gurii i micrilor de lateralitate, constituie o adaptare la staiunea biped, caracteristic omului. La copil articulaia temporo-mandibular este mult mai lax, permind tulburri temporare ale ocluziei dentare. Capul condilului mandibular este mult mai plat, putndu-se realiza micri mai largi de propulsie i retropulsie. Caracteristic articulaiei temporo-mandibulare bogia receptorilor de la acest nivel. Ei sunt mecanoreceptori i nociceptori. Mecanoreceptorii sunt sensibili la tensiunea din ligamentele i capsula articular, ct i la poziia intraarticular a condilului mandibular. Ei pot fi clasificai n receptori de tip I (corpusculi Ruffini) situai superficial n capsula articular, receptori de tip II (corpusculi Pacini) situai profund n capsul, receptori de tip III (corpusculi Golgi) i receptori de tip IV (terminaii nervoase libere). O parte din fibrele receptorilor de tip IV (cele nemielinizate) sunt de fapt nociceptori sau algoreceptori, sensibili la stimulii dureroi din aceast zon. Trebuie menionat c receptorilor de tip III (corpusculii Golgi) li se adaug i fusurile neuromusculare implicate n controlul adecvat al activitii musculare. Ei funcioneaz ca receptori senzoriali, fiind informai continuu asupra gradului de contracie muscular, deci asupra lungimii fibrei musculare. Spre deosebire de fusuri, distribuite n tot corpul muchiului, printre fibrele striate, corpusculii tendinoi Golgi sunt localizai n tendoane, informnd asupra tensiunii musculare. 29
Ca urmare a stimulrii tuturor receptorilor situai n sfera articulaiei temporo-mandibulare i n muchii din aceast regiune, pot fi controlate micrile mandibulei, poziia ei de repaus, reflexele masticatorii i n mai mic msur reflexele salivare, acte fiziologice de importan major i cu o funcionalitate complex. VII. MASTICAIA Reprezint procesul complex prin care alimentele solide i semisolide introduse n cavitatea bucal sunt prelucrate mecanic i impregnate cu saliv n scopul formrii bolului alimentar. Acest proces se realizeaz prin participarea activ a mandibulei, maxilarului superior, dinilor, limbii, obrajilor i buzelor i constituie primul act motor iniiat pe cale reflex sau voluntar la nivelul tractului digestiv. Cicluri masticatorii Masticaia se desfoar sub forma unor succesiuni de cicluri masticatorii ce ncep n momentul prehensiunii alimentelor i se ncheie n momentul deglutiiei. Analiza ciclurilor masticatorii a fost efectuat prin descrierea traiectoriei punctului interincisiv inferior. Acesta descrie elipse succesive revenind ritmic la punctul de plecare, fiecare elips constituind un ciclu masticator. Murphy a distins n cadrul unui ciclu masticator urmtoarele faze: faza de preparare, corespunztoare coborrii punctului incisiv; faza de contact cu bolul alimentar reprezentat de nceputul urcrii mandibulei; faza de strivire, caracterizat prin urcarea punctului incisiv fr a realiza nc un contact interdentar i fazele de mcinare i de ocluzie centrat care ncheie ciclul masticator. Ciclurile masticatorii se succed i n pian frontal: faza preparatoare se efectueaz nspre dreapta sau nspre stnga, ciclul formndu-se n diducie stng sau dreapt. n cursul unui ciclu masticator condilul mandibular activ se deplaseaz i el nainte i n jos iar apoi este dirijat n afar i n sus dup ce punctul interincisiv atinge ocluzia central. Traiectoriile descrise n timpul unui ciclu masticator de ctre punctele interincisiv i condilian depind n primul rnd de anatomia ocluzal individual ct i de caracteristicile fizice ale alimentului mestecat. O influen deosebit asupra masticaiei o exercit educaia i obiceiurile masticatorii precum i suprafaa total a contactelor interdentare n poziia de intercuspidare maxim. S-a constatat c 80% dintre indivizi execut procesul masticator unilateral alternativ, 12% execut cicluri masticatorii pe o singur parte i numai 8% prezint o masticaie bilateral simultan, implicnd mai ales micri de ridicare i coborre ale mandibulei i foarte puine micri de diducie. Eficiena procesului masticator pe lng suprafaa de contact ntre dinii arcadelor n poziie de intercuspidare maxim, depinde i de forele dezvoltate de muchii masticatori. Orice muchi dezvolt o for de contracie proporional cu suprafaa sa de aciune, valoarea sa maxim fiind de 10 kg/cm2. n medie, muchii temporali dezvolt o for de 35 kg/4,5cm2, muchii pterigoidieni externi 28 kg/4 cm2, muchii maseteri 26 kg/3,87 cm2 iar pterigoidienii interni 16 kg/2,3cm2. n cursul micrilor masticatorii asupra dinilor se exercit presiuni verticale cuprinse ntre 30-80kg (n cazul alimentelor solide) i presiuni mai mici ntre 13-15kg pentru alimentele semisolide. Raportnd presiunile exercitate asupra dinilor de ctre bolul alimentar la suprafaa lor ocluzal, se constat c presiunea vertical are valoarea cea mai mare la nivelul dinilor frontali i descrete ctre molarii de 6 ani care au suprafa ocluzal cea mai mare, crescnd apoi uor ctre molarii de minte, ce au o suprafa ocluzal mai redus. Sisteme de prghii Fora dezvoltat de musculatura masticatorie i rezistena opus de alimentele aflate ntre suprafeele triturante, alturi de punctul de sprijin oferit de articulaia temporo-mandibular (condilul pivotant) alctuiesc un sistem de prghii. n micrile de ridicare i coborre ale mandibulei se realizeaz o prghie de ordinul III, n care rezistena este reprezentat de alimentele dintre arcadele dentare, fora este aplicat n locul de inserie al 30
muchilor ridictori ai mandibulei iar punctul de sprijin i constituie articulaiile temporo-mandibulare ce acioneaz simultan, simetric. n micrile de lateralitate ale mandibulei se realizeaz prghii de ordinul II. n acest caz o articulaie temporo-mandibular constituie punctul de sprijin (condilul pivotant), micrile fiind executate n cealalt articulaie, care devine punct de aplicaie al forei; rezistena este reprezentat tot de bolul alimentar plasat ntre aceste dou puncte. Pe msura desfurrii ciclurilor masticatorii i a funcionrii prghiilor temporo-mandibulare, amplitudinea micrilor masticatorii descrete odat cu mcinarea alimentului i impregnarea sa cu saliv. Fiecare ciclu masticator dureaz ntre 0,5-0,7 secunde, succedndu-se n serii iniiate de momentul introducerii alimentelor n gur i ntrerupte n momentul deglutiiei. Mai multe cicluri masticatorii alctuiesc un proces masticator ce se poate desfura ritmic sau aritmie, n funcie de modelul propriu de masticaie al fiecrui individ. Muchii i micrile masticatorii Muchii masticatori constituie componenta activ a procesului de masticaie, asigurnd micrile mandibulei. Micrile de coborre, ridicare, lateralitate, de propulsie i retropulsie sunt realizate prin contraciile succesive ale musculaturii masticatorii principale: - Muchii: maseter, fasciculele anterior i mijlociu ale muchiului temporal, pterigoidianul intern, realizeaz ridicarea mandibulei, crend fora necesar pentru tierea, zdrobirea i mcinarea alimentelor. Ei acioneaz simetric i produc o contracie de tip tetanic a crei for depinde de numrul de uniti motorii n activitate. Sunt inervai de ramura mandibular a nervului trigemen ca i toi ceilali muchi mobilizatori ai mandibulei. - Muchii digastric (pntecele anterior), milohioidianul i geniohioidianul realizeaz micarea de coborre a mandibulei, precedat ns de poziionarea fix a osului hioid, prin contracia muchilor subhioidieni (omohioidianul, tirohioidianul i stemohioidianul). n timp ce digastricul i milohioidianul sunt inervai de fibre din ramura mandibular a trigemenului, muchiul geniohioidian ct i cei subhioidieni sunt inervai de ramuri ale nervului hipoglos. La micarea de coborre a mandibulei, n prghiile de gradul III, contribuie i contracia bilaterala a muchilor pterigoidieni externi. - Contracia alternativ a pterigoidienilor externi alturi de contracia fasciculului posterior al muchiului temporal ipsilateral ct i a muchiului digastric (pntecele anterior), milohioidian i geniohioidian realizeaz micrile de lateralitate ale mandibulei. - Contracia simultan a pterigoidienilor interni i externi realizeaz micrile de propulsie ale mandibulei. - Muchii temporali prin contracia fasciculelor mijlocii concomitent contraciei geniohioidienilor realizeaz retropulsia mandibulei. Musculatura masticatorie nu are o structur omogen i de aceea nici micrile mandibulare nu sunt rezultatul unei succesiuni ritmice de contracii simple ale unui anumit grup muscular, ci ele sunt produse prin contracia mai multor fascicule musculare cu apartenen diferit. Activitii muchilor mobilizatori ai mandibulei li se adaug n procesul masticaiei, contracia muchilor orofaciali, orbicularul buzelor i buccinator, inervai de nervul facial. Ei realizeaz nchiderea orificiului bucal i respectiv adaptarea pereilor laterali ai cavitii bucale la coninutul su. Un alt grup de muchi implicai n desvrirea procesului de masticaie este reprezentat de muchii intrinseci i extrinseci ai limbii inervai de nervul hipoglos i respectiv glosofaringian. Limba, prin musculatura sa, particip la introducerea alimentelor ntre arcadele dentare, la amestecarea alimentelor cu saliva n vederea formrii bolului alimentar i n final la mpingerea sa n faringe. Rolul activ al limbii nu se limiteaz la masticaie, micrile sale fiind importante n actul deglutiiei i fonaiei. Ca urmare a intrrii n activitate a componentei osteo-musculare a cavitii bucale este posibil n procesul de masticaie, prelucrarea mecanic a hranei: 31
- Tierea hranei. Prin contracia fasciculului posterior al temporalului are loc o micare de retropulsie a mandibulei dup care, sub aciunea ridictorilor, are loc propulsia i ridicarea mandibulei, apropiind astfel incisivii i fcnd posibil tierea hranei. n final mandibula revine n poziia de ocluzie centrat i ciclul se reia. n cazul alimentelor cu consisten crescut ridicarea mandibulei este urmat de retropulsie brusc, realizndu-se strivirea lor. - Zdrobirea hranei. Prin contracia muchilor ridictori ai mandibulei este efectuat zdrobirea alimentelor, n principal de ctre premolari, n finalul acestui timp revenindu-se la ocluzia centrat. - Mcinarea hranei. Este realizat prin micri de lateralitate alternative, mai ales de ctre molari. n timpul masticaiei deplasarea alimentelor n gur stimuleaz receptorii bucali, iar senzaiile gustative i olfactive stimuleaz secreia salivar, gastric i pancreatic, cu rol benefic n digestie. Reglarea masticaiei Cercetrile asupra reglrii masticaiei au fost iniiate n 1917 de Sherington pe pisica decerebrat. S-a crezut atunci c masticaia este un act pur reflex. Cercetri ulterioare pe animale i oameni au artat ns c masticaia este un proces parial voluntar, aflndu-se sub controlul sistemului nervos i avnd la baz reflexe necondiionate i condiionate. n realizarea reflexelor masticaiei sunt implicai n primul rnd receptorii din cavitatea bucal: receptori tactili - presionali, termici i dureroi. Presoreceptorii sunt rspndii de asemeni i n parodoniu, fiind sensibili la presiunea exercitat de ctre alimente asupra dintelui n timpul masticaiei. Importani n reglarea masticaiei sunt i receptorii situai n fusurile neuromusculare, n muchii masticatori (receptori Golgi) i n articulaia temporo-mandibular. Toi acetia sunt stimulai de tensiunea creat n timpul masticaiei n muchi, tendoane i capsula articular. I. n momentul introducerii alimentelor, deschiderea cavitii bucale este realizat prin coborrea voluntar a mandibulei, executat sub comanda impulsurilor pornite din aria 4 cortical motorie. De aici, prin fasciculul piramidal cortico-bulbar i apoi prin ramul mandibular al nervului trigemen, impulsurile motorii ajung la pntecele anterior al muchiului digastric, muchii milohioidian, geniohioidian i pterigoidian extern. Prin contracia lor este cobort n mod voluntar mandibula. II. n urma coborrii voluntare a mandibulei i introducerii alimentelor n gur se produce ridicarea reflex a mandibulei prin stimularea fusurilor neuromusculare, a fibrelor anulospirale tip I A, impulsurile pornite de la acest nivel fiind transmise la nucleul senzitiv mezencefalic al trigemenului. De aici impulsurile ajung la nucleul motor al trigemenului din punte, de unde este comandat contracia muchilor maseter, temporal i pterigoidian intern (fasciculul anterior i mijlociu). Astfel, mandibula este ridicat printr-un reflex miotatic nchizndu-se cavitatea bucal. III. Stimularea presoreceptorilor bucali din parodoniu, ligamente periodontale, dentin, mucoasa gingival, mucoasa palatului dur i suprafaa dorsal a limbii, produce impulsuri ce sunt transmise nucleului senzitiv mezencefalic al trigemenului. Dup o ntrziere sinaptic de 1,7 msec, impulsurile ajung la nivelul nucleului motor pontin al trigemenului de unde, prin generarea potenialelor postsinaptice excitatorii va fi comandat coborrea mandibulei. Cnd aceste poteniale ating pragul de excitaie este generat potenialul de aciune care, pe calea nervului mandibular, comand muchilor cobortori ai mandibulei coborrea reflex i deschiderea cavitii bucale. IV. O nou nchidere a cavitii bucale se realizeaz printr-un reflex miotatic, ca urmare a stimulrii fusurilor neuromusculare. Succesiunea de ridicri i coborri ale mandibulei este realizat printr-un lan de reflexe, acestor micri alturndu-li-se i micrile de lateralitate, propulsie i retropulsie mandibular ce concur la prelucrarea mecanic a alimentelor. Actul masticaiei este declanat de introducerea alimentelor n cavitatea bucal, care vor stimula receptorii gustativi, tactili, termici, dureroi i proprioreceptorii de la acest nivel. Stimularea receptorilor 32
gustativi prezeni n cavitatea bucal este de mai mic importan n reglarea masticaiei. Aferentele ce pornesc de la aceti receptori sunt transmise pe calea nervilor facial i glosofaringian, spre deosebire de impulsurile cu importan major n reglarea masticaiei generate de stimularea receptorilor tactili i proprioceptivi ce sunt transmise prin nervii linguali - ramuri ale trigemenului. De la nivelul fusurilor neuromusculare din muchii ridictori ai mandibulei, impulsurile sunt transmise prin prelungirile periferice ale neuronilor pseudounipolari din nucleul mezencefalic trigeminal. Axonii acestor neuroni transmit informaiile nucleului motor pontin trigeminal, la care vin i eferenele motorii corticale de-a lungul tractului piramidal, i de la care pornesc fibre eferente ce comand ridicarea mandibulei. Prelungirile periferice ale neuronilor din nucleul mezencefalic trigeminal preiau informaiile i de la mecanoreceptorii din mucoasa bucal, din structurile parodoniului i din articulaia temporo-mandibular. Spre deosebire de axonii primilor neuroni menionai din nucleul mezencefalic, prelungirile centrale ale acestora din urm, nainte de a ajunge n nucleul motor pontin trigeminal de unde este comandat micarea de coborre a mandibulei, fac sinaps n nucleul supratrigeminal considerat centrul coordonator al masticaiei. Tot spre acest centru sunt trimise prelungirile centrale ale neuronilor din nucleii principali pontini, subnucleul oralis, subnucleul interpolaris i subnucleul caudalis. n aceti patru nudei ajung axonii neuronilor pseudounipolari din ganglionul lui Gasser, ale cror dendrite sunt conectate cu presoreceptorii din mucoasa bucal, parodoniu i articulaia temporo-mandibular, ct i cu termo i algoreceptorii din cavitatea bucal, a cror stimulare n procesul masticaiei este de importan redus la om. Cu excepia fusurilor neuromusculare de la care prin intermediul nucleului mezencefalic al trigemenului impulsurile sunt transmise direct nucleului motor pontin realiznd prin reflex miotatic micarea de ridicare a mandibulei, toi ceilali receptori din structurile cavitii bucale (presoreceptori, termoreceptori, algoreceptori) transmit informaii nucleului supratrigeminal, de la care, n urma conexiunilor cu nucleul motor pontin sunt generate impulsuri motorii efectoare ce realizeaz micarea de coborre a mandibulei. Se observ c reflexul de ridicare a mandibulei este bineuronal, monosinaptic, iar reflexul de coborre a mandibulei este multineuronal polisinaptic, cu neuroni intercalri situai n nucleul supratrigeminal. Complexitatea procesului de masticaie const n faptul c el se afl sub influena altor centri nervoi (cerebel, nuclei bazali, nudei amigdalieni, nudei hipotalamici) ct i sub influene corticale motorii demonstrate de absena masticaiei la nou-nscut i instalarea ei n urma procesului de nvare. S-a demonstrat c i n absena stimulului alimentar poate avea loc o masticaie pur voluntar ceea ce dovedete implicarea cert a cortexului n acest proces. Masticaia este deci un act parial voluntar, realizat prin coordonarea activitii diverselor grupe musculare, aflat sub controlul sistemului nervos i avnd la baz reflexe necondiionate i condiionate. VIII. DEGLUTITIA Deglutiia reprezint mecanismul complex prin care se realizeaz trecerea bolului alimentar din cavitatea bucal n faringe i esofag pn n stomac. Este o funcie permanent, desfurat att n perioada de veghe ct i de somn, n timpul i n afara perioadelor digestive, de minimum 50 de ori n timpul somnului i de minim 200 de ori n timpul meselor. n raport cu segmentele strbtute de bolul alimentar se descriu trei timpi ai deglutiiei: timpul bucal; timpul faringian i timpul esofagian. Timpul bucal este voluntar i const n aezarea bolului alimentar pe faa dorsal a limbii care l preseaz, ca un piston, pe bolta palatin napoia incisivilor superiori. Muchii intrinseci dau limbii forma de jgheab, n care se muleaz bolul alimentar; muchii geniohioidieni trag puin limba nainte pentru a da libertate vrfului limbii s se etaleze i s se rsfrng napoi spre bolta palatin.
33
Muchii milohioidieni se contract i ridic planeul bucal odat cu limba, care va apsa cu toat fora pe bolta palatin i n poriunea posterioar, unde se declaneaz, n mod reflex, secreia glandelor mucoase, fcnd mai alunecos bolul alimentar. Limba exercit pe bolta palatin o presiune cuprins ntre 41 i 70 g/cm2. Ea este mai important pe prile laterale ale boltei i variaz cu consistena alimentelor; cea mai redus presiune o determin nghiirea lichidelor. Exercitarea unei presiuni excesive n partea posterioar a boltei palatine determin vestibulizarea dinilor. Lichidele se beau prin suciune sau aspiraie, datorit presiunii negative realizat n cavitatea bucal, prin retracia limbii, fr obturaia limb-palat. Cnd cavitatea bucal se umple cu lichid, partea posterioar a limbii coboar, maxilarul inferior se ridic, nchide cavitatea i, printr-o micare de baleiaj a limbii, lichidele sunt mpinse n faringe. La copilul sugar deglutiia este realizat prin micri peristaltice ale buccinatoruiui i constrictorului superior al faringelui, fr a fi nevoie de ridicarea mandibulei i contactul ntre arcadele alveolare. Lichidul nghiit coboar de-a lungul valeculelor - anuri adnci pe faa posterioar a bazei limbii direct spre esofag, fr a fi necesar nchiderea laringelui i deci ridicarea mandibulei la fiecare nghiitur, limba rmnnd interpus ntre arcade. La introducerea alimentaiei mixte a sugarului, se reduc micrile de propulsie i retropulsie ale mandibulei. Buzele i buccinatorii se contract peristaltic numai n timpul suptului. Dup vrsta de un an, cnd copilul nu va mai suge, aciunea sfincterian a buzelor i buccinatoruiui este compensat prin ridicarea mandibulei i contactul dintre dini la fiecare nghiitur. Limba nu se mai interpune ntre dini i joac rolul de transportor al bolului alimentar spre faringe. Vrful limbii etalat, apas pe regiunea palatului anterior, n dreptul gurii nazopalatine, ca un piston. Se trece astfel de la deglutiia infantil la cea adult. Meninerea prelungit a deglutiiei infantile cu apsarea dinilor frontali de ctre limb produce vestibulizarea dinilor frontali (pro-alveolodenia), inocluzie vertical i strmtorarea arcadelor alveolodentare i pe de alt parte palatinizarea frontalilor. Timpul bucal al deglutiiei este scurt, ntre 0,2-0,3 secunde pn la 1,2 secunde n raport cu natura, coninutului alimentar nghiit. Timpul faringian este un timp involuntar, realizat n apnee, ncepnd n momentul n care bolul alimentar atinge anumite pri ale mucoasei faringiene i declaneaz reflex micrile tipice de la nivelul celor patru orificii din rspntia faringian. Prin relaxarea muchilor palatogloi i faringopalatini, care ndeprteaz stlpii anteriori i cei posteriori, este permis trecerea boiului alimentar. Imediat dup aceasta, se produce nchiderea istmului buco-faringian prin contracia acelorai muchi i prin apropierea amigdalelor. Orificiul nazo-faringian se nchide prin ridicarea vlului palatin ca urmare a contraciei muchilor ridictori i tensori ai vlului palatin. n acelai timp, se contract i muchiul salpingo-faringian, nchizndu-se astfel orificiul trompei lui Eustachio. Orificiul laringo-faringian se nchide prin ridicarea hioidului, mandibulei, limbii, laringelui i prin coborrea epiglotei, ct i prin apropierea corzilor vocale ca urmare a contraciei muchilor aritenoizi. Muchii implicai n realizarea acestei etape sunt: muchii ridictori ai mandibulei, muchii suprahioidieni (milohioidian i geniohioidian), muchii tirohioidieni, muchii stilogloi i hiogloi. Rentoarcerea bolului alimentar n cavitatea bucal n aceasta faz a deglutiiei, nu este posibil, din cauza meninerii contraciei musculaturii ce a determinat mpingerea bolului n faringe, contraciei muchilor faringo-palatini i gloso-palatini care apropie stlpii faringelui, micornd astfel diametrul istmului buco-faringian. Etapa faringian a deglutiiei este cea mai scurt (0,1 - 0,2 sec.) i cea mai complex, necesitnd un fin control al reflexelor nchise ntre receptorii buco-faringieni i centrul deglutiiei. 34
Prin contracia muchilor intrinseci i extrinseci faringieni, bolul alimentar progreseaz n faringe, ca urmare a undei peristaltice ce creeaz o presiune de 70-100 cm H2O, asigurnd naintarea sa. Viteza de deplasare a bolului alimentar, studiat cu metode cinefluoroscopice este diferit n raport cu segmentele strbtute. De la o vitez de 3 m/sec, la intrarea n faringe, se ajunge la 9 m/sec, apoi descrete la 4 m/s, pentru ca la intrarea n esofag s fie sub 2 m/s. Timpul esofagian al deglutiiei este un timp involuntar ce const n trecerea bolului alimentar prin esofag, cu viteze diferite n funcie de consistenta sa. Lichidele l strbat rapid, acumulndu-se de obicei deasupra cardiei pn la deschiderea acestui sfincter de ctre unda peristaltic declanat n momentul trecerii prin esofag. Solurile alimentare solide strbat mai lent esofagul, n aproximativ 5-6 secunde, cu vitez mai mare n esofagul proximal, parcurs n 1,5-2 secunde, datorit prezenei musculaturii striate la acest nivel i mult mai lent, n aprox. 3 sec, n esofagul distal, ce prezint musculatur neted. Undele peristaltice ce asigur progresia bolului prin esofag sunt de trei tipuri: unde primare, declanate de excitarea receptorilor bucofaringieni, aflate sub controlul inervaiei extrinseci esofagiene; unde secundare, declanate de excitarea receptorilor mucoasei esofagiene din dreptul arcului aortic. n urma stimulrii acestor receptori de ctre distensia mecanic local a musculaturii netede din cele 2/3 inferioare ale esofagului, sau de ctre iritaiile locale produse de aciditatea refluxului gastroesofagian, sunt generate undele secundare, care alturi de cele primare, prin propagarea lor, asigur evacuarea coninutului esofagian; undele teriare, apar spontan n regiunea inferioar a esofagului, sunt contracii locale, neregulate, asimetrice, frecvent ntlnite la btrni i sunt coordonate de inervaia intrinsec a esofagului distal. Prezena lor la adultul tnr nu este fiziologic, putnd indica existena proceselor proliferative esofagiene. Trecerea bolului alimentar n stomac prin sfincterul cardial este condiionat de existena unei presiuni intraesofagiene cuprins ntre 2-7 cm H2O, ce va determina deschiderea cardiei i ptrunderea alimentelor n stomac. Uneori, la aproximativ 20-30 minute dup mas se pot produce n esofag regurgitri de coninut gastric, contracarate imediat de unde peristaltice cu propagare distal ce mping coninutul regurgitat din nou n stomac. n perioadele interdigestive, sfincterul cardial este nchis, acesta putndu-se deschide doar dac presiunea intragastric depete valoarea de 25 cm H2O. Reglarea deglutiiei Este realizat prin mecanisme n principal nervoase i secundar umorale (la nivelul sfincterului esofagian distal). Stimulii care declaneaz reflexul deglutiiei acioneaz asupra mecanoreceptorilor, chemoreceptorilor i termoreceptorilor, situai n mucoasa de la baza limbii, din vlul palatin, din stlpii anteriori ai faringelui, din regiunea faringo-amigdalian (zona reflexogen Wasilieff). De la nivelul acestor zone reflexogene pornesc cile aferente, incluse n ramura mandibular a trigemenului, nervul glosofaringian, nervul laringeu superior, nervul recurent i ramurile esofagiene ale nervului vag. Pe aceste ci, stimulii ajung n nucleii senzitivi ai nervilor cranieni corespunztori i la centrul deglutiiei situat n substana reticulat bulbar, n planeul ventriculului IV. Fibrele eferente care pornesc de la acest centru transmit impulsuri ctre nucleii motori ai nervilor cranieni V, IX, X i XII. De la aceti nudei pornesc eferente motorii pentru muchii mobilizatori ai mandibulei (prin nervul V - trigemen); pentru musculatura limbii (prin nervul XII - hipoglos); pentru musculatura limbii i a faringelui (prin nervul IX - glosofaringian) i pentru musculatura faringoesofagian prin nervii X - vagi. Tot vagul este cel care controleaz activitatea sfincterului cardial. 35
Controlul umoral se realizeaz prin intermediul gastrinei, motilinei, colecistochininei, secretinei, glucagonului, prostaglandinelor, VIP, influena lor manifestndu-se la nivelul sfincterului esofagian distal. IX. FONAIA Laringele este considerat organul fonator propriu-zis, la nivelul su fiind produse sunetele, prin vibraia corzilor vocale, odat cu trecerea aerului expirat sub presiune, prin orificiul glotic. Corzile vocale sunt alctuite din muchii tiroaritenoidieni, esut membranos i ligamentul vocal, aflndu-se ntre cartilajul tiroid (anterior),cartilajele aritenoide (posterior) i cartilajul cricoid (la baz). Prin contracia ritmic a muchiului vocal al corzilor, sunt imprimate micri vibratorii coloanei de aer, dnd natere sunetului laringian. Contracia ritmic a acestor muchi i a celorlali muchi laringieni se realizeaz printr-un reflex, care se nchide la nivelul centrilor corticali i subcorticali ai fonaiei. Aceti centrii nervoi au legturi strnse cu centrii vorbirii i auzului. Prima etap n mecanismul producerii de sunete const n pregtirea pentru expir, timp n care glota st nchis sau uor ntredeschis, crescnd astfel presiunea subglotic. n mod normal, n vocea de conversaie, valorile presiunii subglotice variaz ntre 4 i 6 cm H2O putnd ajunge la peste 20 cm H2O la cntreii de oper. Aceast presiune a coloanei de aer stimuleaz proprio receptorii din muchiul vocal i determin mpingerea lateral a corzilor vocale. Drept urmare, aerul trece prin glota seminchis spre cavitatea nazofaringian, producnd vibraiile corzilor vocale. n urma trecerii coloanei de aer se creeaz un vacuum parial ntre corzile vocale, din cauza fenomenului BERNOULU, glota este renchis i odat cu creterea presiunii subglotice, rencepe vibraia corzilor vocale. Caracteristicile sunetelor laringiene emise variaz n funcie de presiunea coloanei de aer, lungimea i tensiunea corziior vocale. Rolul cavitii bucale n fonaie mpreun cu vestibulul laringian, faringele, cavitatea nazal i chiar cavitatea toracic, spaiul cavitii bucale alctuiete aparatul de rezonan al fonaiei, cu rol de amplificator al tonului vocal laringian. La nivelul laringelui se produc sunete nearticulate. Acestea devin articulate, formnd vocea uman, la nivelul cavitilor de rezonan superioare: cavitatea bucal, faringele i cavitile nazale. Un rol deosebit n emiterea sunetelor articulate revine cavitii bucale prin poziia i forma limbii, poziia buzelor, obrajilor, a palatului moale i a mandibulei. Toate aceste formaiuni se mobilizeaz n timpul fonaiei pentru a produce un anumit sunet. Caracteristicile sunetelor depind nu de sunetele produse n laringe, ci de modificrile de volum ale camerelor de rezonan la nivelul crora se produce vocea, emindu-se vocalele i consoanele (foneme). Prin ngustarea unor spaii laringo-faringiene n momentul trecerii sunetului fundamental, nearticulat, se pronun vocalele. Pe de alt parte, prin ntreruperea trecerii aerului prin faringe sau cavitatea bucal, coloana de aer a sunetelor emise de laringe sufer ntreruperi n diferite regiuni ale bucofaringelui producnd astfel diferite consoane. Dup locul ntreruperii coloanei de aer, consoanele se pot clasifica n: bilobiale ("b", "p"), labiodentare ("f", "v"), dentolinguaie ("d", "t"), linguopalatine sau faringiene ("g", "k"), nazale ("m", "n"), linguale ("i", "r") etc. Fonaia n ansamblul ei este un proces ce se nva i se desvrete lent, n civa ani. n funcie de gradul individual de instruire, acest fenomen poate fi modulat la nivelul cavitilor bucofaringiene, cptnd inflexiuni plcute auzului. Deoarece la procesul articulrii sunetelor particip mai multe formaiuni (limb, buze, palatul moale), acestea trebuie s fie riguros controlate de cortexul motor, pentru a evita diferenele de pronunie. Reglarea fonaiei se face deci pe cale nervoas, n lobul prefrontal ar exista centrul vorbirii, care prezint conexiuni importante cu centrii respiratori, realiznd astfel coordonarea absolut necesar ntre fonaie i respiraie. De altfel, att laringele ct i bucofaringele sunt organe cu funcie dubl, respiratorie i fonatorie. 36
De asemeni sunt stabilite conexiuni i cu centrii auditivi i vizuali din lobii temporal i respectiv occipital realizndu-se astfel circuite integratoare auditiv-vizuale. n reglarea nervoas a fonaiei intervine i sistemul motor extrapiramidal ce asigur controlul activitii motorii la nivelul structurilor bucofaringiene. Distrugerea centrului cortical al fonaiei poate duce la anartrie, fr a fi nsoit ns de tulburri ale deglutiiei, masticaiei sau respiraiei (centrii coneci dar cu dubl reprezentare cortical). Perturbri ale fonaiei se pot produce i n cazul leziunilor i malformaiilor organelor fonatorii (buze, limb, palat moale) ct i n deformri dentomaxilare. Acestea pot fi obiectivate prin: electrologografie care nregistreaz simultan cuvintele i vibraiile corzilor vocale; fonografie ce d indicaii asupra amplitudinii, frecvenei, timbrului i duratei sunetelor; glotografie ce apreciaz micrile corzilor vocale. Cu ajutorul acestor metode se poate pune diagnosticul etiologic al tulburrii de fonaie i se poate aprecia evoluia afeciunii n timpul terapiei.
37
S-ar putea să vă placă și
- Cavitatea BucalaDocument48 paginiCavitatea BucalaRoxana Azamfiroaei100% (1)
- Aparatul DigestivDocument10 paginiAparatul DigestivPatricia AdamÎncă nu există evaluări
- Ifrim PDFDocument263 paginiIfrim PDFDaniAlbÎncă nu există evaluări
- Partea I Curs IfrimDocument57 paginiPartea I Curs IfrimMarius Petru BocorÎncă nu există evaluări
- ANATOMIADocument2 paginiANATOMIADaiana ZaicicoÎncă nu există evaluări
- Anatomia Şi Fiziologia Cavităţii BucaleDocument19 paginiAnatomia Şi Fiziologia Cavităţii BucaleMarian Costin Radulescu50% (2)
- 7 - Anatom TopDocument5 pagini7 - Anatom TopCristina BeşliuÎncă nu există evaluări
- HistologieDocument39 paginiHistologieConstantin UntilaÎncă nu există evaluări
- Cavitatea BucalaDocument10 paginiCavitatea BucalaDiana BeatriceÎncă nu există evaluări
- 2 AnatomieDocument73 pagini2 AnatomiecorinabeseleaÎncă nu există evaluări
- A. Morfologia Normala A Mucoasei OraleDocument101 paginiA. Morfologia Normala A Mucoasei OraleAli ȘerbănescuÎncă nu există evaluări
- Aparatul DigestivDocument3 paginiAparatul DigestivMarian DeliuÎncă nu există evaluări
- Mucoasa Cavit - BucalDocument5 paginiMucoasa Cavit - BucalRadu CraciunÎncă nu există evaluări
- Cavitatea Bucala-Curs 4Document3 paginiCavitatea Bucala-Curs 4Nosaci AndreiÎncă nu există evaluări
- Anatomie 2Document151 paginiAnatomie 2Andreea OlteanÎncă nu există evaluări
- Aparat Digestiv Medicina VeterinaraDocument95 paginiAparat Digestiv Medicina Veterinaramihai2242Încă nu există evaluări
- Cap. 2 Aparatul Digestiv-HistologieDocument21 paginiCap. 2 Aparatul Digestiv-HistologieBuhas MadalinaÎncă nu există evaluări
- Patologie Orala Monea Adriana Maria Note de Curs 2015Document148 paginiPatologie Orala Monea Adriana Maria Note de Curs 2015Maria Cristina ChelaruÎncă nu există evaluări
- Răspunsuri Elaborări Metodice Morfologia SsDocument35 paginiRăspunsuri Elaborări Metodice Morfologia SsCharlie McdonnellÎncă nu există evaluări
- Curs 6 - Cavitatea BucalaDocument13 paginiCurs 6 - Cavitatea BucalaermilovelenaÎncă nu există evaluări
- MorfologiaDocument25 paginiMorfologiatatiana0ostahii100% (1)
- Cavitas OrisDocument10 paginiCavitas OrisSmarandescu Alexandru IonutÎncă nu există evaluări
- AdmDocument27 paginiAdmSerg SgÎncă nu există evaluări
- Cavitatea Bucala TMDocument64 paginiCavitatea Bucala TMAndreea MitranÎncă nu există evaluări
- Lucrarea Practic (Nr. 8 - Ap - Dig. PrediafragmaticDocument10 paginiLucrarea Practic (Nr. 8 - Ap - Dig. PrediafragmaticIoana-camelia StanilaÎncă nu există evaluări
- Anatomie BucalaDocument36 paginiAnatomie BucaladolaÎncă nu există evaluări
- Histologie Curs 4Document7 paginiHistologie Curs 4Bianca IonițăÎncă nu există evaluări
- Splanchnologie IasiDocument351 paginiSplanchnologie IasiB.L.C.SÎncă nu există evaluări
- CAP1Document136 paginiCAP1Schidu DanielÎncă nu există evaluări
- Aparat Digestiv - Pe ScurtDocument33 paginiAparat Digestiv - Pe ScurtCotovici Catana AlinaÎncă nu există evaluări
- Cavitatea BucalaDocument53 paginiCavitatea BucalaDiana PitisÎncă nu există evaluări
- Anato CursDocument8 paginiAnato CursCarmen CmÎncă nu există evaluări
- Anatomia Aparatului DigestivDocument21 paginiAnatomia Aparatului DigestivSandu Ionut RobertÎncă nu există evaluări
- Patologie Tot VERIFICATDocument157 paginiPatologie Tot VERIFICATSalwa SellyÎncă nu există evaluări
- PatologieDocument118 paginiPatologieMariaÎncă nu există evaluări
- Anatomie SesiuneDocument60 paginiAnatomie SesiuneEduard.Încă nu există evaluări
- Aparatul Digestiv Este Alcătuit Din Totalitatea Organelor Care Îndeplinesc Importanta Funcţie de Digestie Şi Absorbţie A Alimentelo1Document297 paginiAparatul Digestiv Este Alcătuit Din Totalitatea Organelor Care Îndeplinesc Importanta Funcţie de Digestie Şi Absorbţie A Alimentelo1auras27mateiÎncă nu există evaluări
- Curs 7 Cavitatea BucalăDocument20 paginiCurs 7 Cavitatea BucalăLumy100% (1)
- Patologie Orala-C 1Document10 paginiPatologie Orala-C 1paulaandreea20Încă nu există evaluări
- BiologieDocument20 paginiBiologiePaula CristianaÎncă nu există evaluări
- Anatomia Si Fiziologia Cavităţii BucaleDocument17 paginiAnatomia Si Fiziologia Cavităţii BucaleTanase Antonel100% (1)
- c1 Particularitati Morfo Functionale Ale Cav BucaleDocument21 paginic1 Particularitati Morfo Functionale Ale Cav BucaleAnamariaMntÎncă nu există evaluări
- Compartimentul Fetei AnterioareDocument7 paginiCompartimentul Fetei AnterioareIrina StratilaÎncă nu există evaluări
- Sistem DigestivDocument15 paginiSistem Digestivcamelia smyÎncă nu există evaluări
- Splanhnologiacurs Anul I Sem IIDocument248 paginiSplanhnologiacurs Anul I Sem IIAnna AnnÎncă nu există evaluări
- Cavitatea BucalaDocument12 paginiCavitatea BucalaIuliana BicecÎncă nu există evaluări
- SUBIECTE REZOLVATE PATOLOGIE ORALA - Final-De TottotDocument90 paginiSUBIECTE REZOLVATE PATOLOGIE ORALA - Final-De TottotAlexandru VladÎncă nu există evaluări
- Elementele Componente Ale Campului Protetic Edentat TotalDocument5 paginiElementele Componente Ale Campului Protetic Edentat TotalNadia Balaescul100% (2)
- (Bio) Anatomia Cavitatii BucaleDocument4 pagini(Bio) Anatomia Cavitatii BucaleDenis BogdanÎncă nu există evaluări
- Cavitatea BucalaDocument12 paginiCavitatea BucalaDaniel Alexandru ParauÎncă nu există evaluări
- Aparatul DigestivDocument139 paginiAparatul DigestivMaria100% (1)
- 5 Sistemul DigestivDocument23 pagini5 Sistemul DigestivzennmotoÎncă nu există evaluări
- AnatomieDocument12 paginiAnatomieIacob Madalina IonelaÎncă nu există evaluări
- LP 7 Esofagul Și StomaculDocument58 paginiLP 7 Esofagul Și StomaculIonela VioletaÎncă nu există evaluări
- Procesul Nursing Al Pacientului Cu Suferinţă Digestivă, În AmbulatoriuDocument9 paginiProcesul Nursing Al Pacientului Cu Suferinţă Digestivă, În Ambulatoriuana12092Încă nu există evaluări
- Fit Generation: Primul Manual De Fitness Din RomaniaDe la EverandFit Generation: Primul Manual De Fitness Din RomaniaEvaluare: 4 din 5 stele4/5 (8)
- Analiza Campaniei PrezidentialeDocument1 paginăAnaliza Campaniei PrezidentialeGigel CovaljievÎncă nu există evaluări
- Analiza Campaniei PrezidentialeDocument2 paginiAnaliza Campaniei PrezidentialeGigel CovaljievÎncă nu există evaluări
- Analiza Campaniei PrezidentialeDocument3 paginiAnaliza Campaniei PrezidentialeGigel CovaljievÎncă nu există evaluări
- Analiza Campaniei PrezidentialeDocument2 paginiAnaliza Campaniei PrezidentialeGigel CovaljievÎncă nu există evaluări
- Analiza Campaniei PrezidentialeDocument2 paginiAnaliza Campaniei PrezidentialeGigel CovaljievÎncă nu există evaluări
- Analiza Campaniei PrezidentialeDocument1 paginăAnaliza Campaniei PrezidentialeGigel CovaljievÎncă nu există evaluări
- Analiza Campaniei PrezidentialeDocument1 paginăAnaliza Campaniei PrezidentialeGigel CovaljievÎncă nu există evaluări
- Analiza Campaniei PrezidentialeDocument3 paginiAnaliza Campaniei PrezidentialeGigel CovaljievÎncă nu există evaluări
- Analiza Campaniei PrezidentialeDocument4 paginiAnaliza Campaniei PrezidentialeGigel CovaljievÎncă nu există evaluări
- IvaDocument9 paginiIvaGigel CovaljievÎncă nu există evaluări
- Analiza Campaniei PrezidentialeDocument3 paginiAnaliza Campaniei PrezidentialeGigel CovaljievÎncă nu există evaluări
- Analiza Campaniei PrezidentialeDocument5 paginiAnaliza Campaniei PrezidentialeGigel CovaljievÎncă nu există evaluări
- Analiza Campaniei PrezidentialeDocument4 paginiAnaliza Campaniei PrezidentialeGigel CovaljievÎncă nu există evaluări
- AlegeriDocument4 paginiAlegeriGigel CovaljievÎncă nu există evaluări
- Experimentul StandofrdDocument7 paginiExperimentul StandofrdGigel CovaljievÎncă nu există evaluări
- Metode de Cercetare SPDocument62 paginiMetode de Cercetare SPMariscaÎncă nu există evaluări
- Institutii DecizionaleDocument62 paginiInstitutii DecizionaleMariscaÎncă nu există evaluări
- 3 4 12 20 Rezolvare Subiecte Sisteme Politice ComparateDocument15 pagini3 4 12 20 Rezolvare Subiecte Sisteme Politice ComparateGigel CovaljievÎncă nu există evaluări
- Vot PDFDocument6 paginiVot PDFGigel CovaljievÎncă nu există evaluări
- Curs de Comunicare PoliticăDocument34 paginiCurs de Comunicare PoliticăMariscaÎncă nu există evaluări
- Aplicatie HE StandardDocument1 paginăAplicatie HE StandardGigel CovaljievÎncă nu există evaluări
- Republica MoldovaDocument12 paginiRepublica MoldovaGigel CovaljievÎncă nu există evaluări
- Suport GuvernantaDocument64 paginiSuport Guvernantaadina_cristiana_1Încă nu există evaluări
- Mecanismul Votului (Rezumat)Document2 paginiMecanismul Votului (Rezumat)Probocis AlexÎncă nu există evaluări
- AlegeriDocument4 paginiAlegeriGigel CovaljievÎncă nu există evaluări
- Constituirea Si Activitatea Partidului National LiberalDocument13 paginiConstituirea Si Activitatea Partidului National LiberalGigel CovaljievÎncă nu există evaluări
- AlegeriDocument4 paginiAlegeriGigel CovaljievÎncă nu există evaluări
- RECENZIEDocument3 paginiRECENZIEGigel CovaljievÎncă nu există evaluări
- Recenzie - Mecanismul VotuluiDocument8 paginiRecenzie - Mecanismul VotuluigerminareÎncă nu există evaluări
- Experimentul StandofrdDocument5 paginiExperimentul StandofrdGigel CovaljievÎncă nu există evaluări