S-ar putea să vă placă și
- Instrumentarul ChirurgicalDocument9 paginiInstrumentarul ChirurgicalLeonard D100% (22)
- Clasificarea Fibrelor NervoaseDocument4 paginiClasificarea Fibrelor NervoaseAlina Simona TeodoroviciÎncă nu există evaluări
- Clasificarea Articulatiilor PDFDocument3 paginiClasificarea Articulatiilor PDFAndreea Diana DrăgoiÎncă nu există evaluări
- Fiziologie Anul 1 Curs 1 DavilaDocument71 paginiFiziologie Anul 1 Curs 1 DavilaTeodor-Nicolae Maciu-Popescu0% (1)
- Polaritatea Tesutului EpitelialDocument7 paginiPolaritatea Tesutului EpitelialAriela SăvuţÎncă nu există evaluări
- MetabolismulDocument144 paginiMetabolismulDumitru NucaÎncă nu există evaluări
- 5p. Compuși MacroergiciDocument5 pagini5p. Compuși MacroergiciMădălina Stănășel50% (2)
- Carte Biochimie Stoian Irina 2Document116 paginiCarte Biochimie Stoian Irina 2Irina StoianÎncă nu există evaluări
- Procese Redox in Organismul UmanDocument7 paginiProcese Redox in Organismul UmanDanielPetreÎncă nu există evaluări
- Biosinteza ProteinelorDocument21 paginiBiosinteza ProteinelorAlice DouglasÎncă nu există evaluări
- Farmacologia Sistemului Nervos VegetativDocument42 paginiFarmacologia Sistemului Nervos VegetativAlice DouglasÎncă nu există evaluări
- Prezentare de Caz Clinic - Semiologie ChirurgicalăDocument7 paginiPrezentare de Caz Clinic - Semiologie ChirurgicalăAlice DouglasÎncă nu există evaluări
- Functiile Maduvei, Functiile NeuronuluiDocument1 paginăFunctiile Maduvei, Functiile NeuronuluilavybbyÎncă nu există evaluări
- Subiecte Rezolvate ColocviuDocument18 paginiSubiecte Rezolvate ColocviuCristina IoanaÎncă nu există evaluări
- ARTICULATIILEDocument4 paginiARTICULATIILETepeş-Stroia EugenÎncă nu există evaluări
- Circulatia ArterialaDocument7 paginiCirculatia ArterialadanielardlscÎncă nu există evaluări
- Muşchii Gatului-Sternocleidomastoidian, Scaleni, Infrahiodieni+M.PrevertebraliDocument3 paginiMuşchii Gatului-Sternocleidomastoidian, Scaleni, Infrahiodieni+M.PrevertebraliBogdan100% (1)
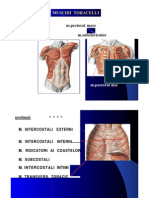
- Muschii ToraceluiDocument7 paginiMuschii ToraceluiBenny ScorpyoÎncă nu există evaluări
- Transmiterea Sinaptica Prin Sinapsele Chimice PDFDocument5 paginiTransmiterea Sinaptica Prin Sinapsele Chimice PDFMădălina NăstuţăÎncă nu există evaluări
- Maduva SpinariiDocument4 paginiMaduva SpinariitudortudorgÎncă nu există evaluări
- Calea PentozofosfațilorDocument7 paginiCalea PentozofosfațilorAlexandra MagaleasÎncă nu există evaluări
- Nervii FesieriDocument3 paginiNervii FesieriDan100% (1)
- Oasele Carpiene PresentationDocument32 paginiOasele Carpiene PresentationBogdan Alexandru CristeiÎncă nu există evaluări
- Plex SacralDocument5 paginiPlex Sacrallexamara100% (2)
- Regiunea Genunchiului Si Gambei Covali IonDocument53 paginiRegiunea Genunchiului Si Gambei Covali IonOctavianÎncă nu există evaluări
- Anatomie Pe Viu Sem 1Document20 paginiAnatomie Pe Viu Sem 1Berladean Natalia100% (2)
- Alcătuirea Generală A Corpului Uman - Sistemul NervosDocument32 paginiAlcătuirea Generală A Corpului Uman - Sistemul NervosEllyzaEllyÎncă nu există evaluări
- 8 Circuite Neuronale 2Document1 pagină8 Circuite Neuronale 2Alex Cu TridentÎncă nu există evaluări
- Sistemul MuscularDocument9 paginiSistemul MuscularDiana AndreeaÎncă nu există evaluări
- ExcitabilitateaDocument27 paginiExcitabilitateastrengara100% (2)
- Nervii SpinaliDocument3 paginiNervii SpinaliDanutz MariusÎncă nu există evaluări
- UroteliulDocument1 paginăUroteliulatd999Încă nu există evaluări
- ReflexeDocument31 paginiReflexeCristina DiaconuÎncă nu există evaluări
- Artera BrahialaDocument1 paginăArtera BrahialaMihaiR24Încă nu există evaluări
- Contractia Musculaturii Muschiului CardiacDocument4 paginiContractia Musculaturii Muschiului CardiacMitrache AureliaÎncă nu există evaluări
- Plexul Cervical DocumentDocument2 paginiPlexul Cervical DocumentTeona MelinteÎncă nu există evaluări
- Totalizare Anatomie 3Document17 paginiTotalizare Anatomie 3Anonymous ck0aGAqZl100% (2)
- Plexul CervicalDocument4 paginiPlexul CervicalAdriana Smerea50% (2)
- Topografia CoapseiDocument3 paginiTopografia CoapseiVictor JuncuÎncă nu există evaluări
- Organizarea Corpului UmanDocument4 paginiOrganizarea Corpului UmanIuliaNaTuFaruÎncă nu există evaluări
- Muschii Anterolaterali Ai ToraceluiDocument2 paginiMuschii Anterolaterali Ai ToraceluiElena-Dana Oprea100% (3)
- Aparatul GolgiDocument3 paginiAparatul Golgipisimiau070% (1)
- Fiziologia MuschilorDocument10 paginiFiziologia MuschilorNicolae Elena Andreea100% (2)
- Nervul FemuralDocument17 paginiNervul FemuralCocosul Cocosului CocosaruluiÎncă nu există evaluări
- ArticulatiiDocument29 paginiArticulatiiAlinaCodreanuÎncă nu există evaluări
- Anul 1 - Fiziologie Grile TipDocument70 paginiAnul 1 - Fiziologie Grile TipIonuț-Ciprian TrancăÎncă nu există evaluări
- CordulDocument8 paginiCordulGuțaÎncă nu există evaluări
- MuschiDocument11 paginiMuschiDumitru MihaiÎncă nu există evaluări
- Fiziologia MuschilorDocument15 paginiFiziologia Muschilorpstefan18100% (1)
- Curburile În Plan SagitalDocument3 paginiCurburile În Plan SagitalDenisaÎncă nu există evaluări
- Analizatorul CutanatDocument16 paginiAnalizatorul Cutanatpaun_alexandra89Încă nu există evaluări
- Articulatiile Membrului Superior LiberDocument3 paginiArticulatiile Membrului Superior LiberPuppa AlexandraÎncă nu există evaluări
- CLASIFICAREDocument18 paginiCLASIFICAREMihai CimpocaÎncă nu există evaluări
- Resp6 2013 PDFDocument44 paginiResp6 2013 PDFMihai Chirila0% (1)
- Muschii BratuluiDocument8 paginiMuschii BratuluiMihai RacovițaÎncă nu există evaluări
- Articulatiile Coloanei Si ToraceluiDocument24 paginiArticulatiile Coloanei Si ToraceluiAlina JigaranuÎncă nu există evaluări
- Bolta PicioruluiDocument2 paginiBolta PicioruluiKitty CatyÎncă nu există evaluări
- Tabel Mușchi Anatomie Stratenco SergiuDocument56 paginiTabel Mușchi Anatomie Stratenco SergiuSergiu StratencoÎncă nu există evaluări
- Transportul ActivDocument8 paginiTransportul ActivElise SarahÎncă nu există evaluări
- ATPDocument2 paginiATPMihaelaFlorentinaÎncă nu există evaluări
- Biochimie II 4Document10 paginiBiochimie II 4ansa110Încă nu există evaluări
- MetabolismulDocument6 paginiMetabolismuleugeniu222Încă nu există evaluări
- Enzime 1Document38 paginiEnzime 1Elena Alina TÎncă nu există evaluări
- Integrarea Metabolismului 1Document25 paginiIntegrarea Metabolismului 1Andreea MayaÎncă nu există evaluări
- BiochimieDocument27 paginiBiochimieAlice DouglasÎncă nu există evaluări
- Ciclul Acizilor Tricarboxilici Şi Calea FosfogluconatuluiDocument20 paginiCiclul Acizilor Tricarboxilici Şi Calea FosfogluconatuluiAlice DouglasÎncă nu există evaluări
- ProteineDocument51 paginiProteineAlice DouglasÎncă nu există evaluări
- HormoniiDocument33 paginiHormoniiAlice DouglasÎncă nu există evaluări
- Metablismul NucleotidelorDocument60 paginiMetablismul NucleotidelorAlice DouglasÎncă nu există evaluări
- Ciclul Acizilor Tricarboxilici Şi Calea FosfogluconatuluiDocument20 paginiCiclul Acizilor Tricarboxilici Şi Calea FosfogluconatuluiAlice DouglasÎncă nu există evaluări
- Glucide (Hidraţi de Carbon)Document46 paginiGlucide (Hidraţi de Carbon)Alice DouglasÎncă nu există evaluări
- Transportul de ElectroniDocument20 paginiTransportul de ElectroniAlice DouglasÎncă nu există evaluări
- Catabolismul ProteinelorDocument84 paginiCatabolismul ProteinelorAlice Douglas100% (2)
- Lipide, Structură Și ProprietățiDocument95 paginiLipide, Structură Și ProprietățiAlice Douglas100% (1)
- Metablismul NucleotidelorDocument60 paginiMetablismul NucleotidelorAlice DouglasÎncă nu există evaluări
- Celula - BiochimieDocument6 paginiCelula - BiochimieAlice DouglasÎncă nu există evaluări
- Introducere Curs Biochimie ClinicăDocument6 paginiIntroducere Curs Biochimie ClinicăAlice DouglasÎncă nu există evaluări
- Reacții Adverse La Medicamente - FarmacologieDocument2 paginiReacții Adverse La Medicamente - FarmacologieAlice DouglasÎncă nu există evaluări
- Anticanceroase - Antineoplazice - FarmacologieDocument26 paginiAnticanceroase - Antineoplazice - FarmacologieAlice DouglasÎncă nu există evaluări
- FarmacotoxicologiaDocument1 paginăFarmacotoxicologiaAlice DouglasÎncă nu există evaluări
- Biomoleculele Și Metode BiochimiceDocument4 paginiBiomoleculele Și Metode BiochimiceAlice DouglasÎncă nu există evaluări
- Farmacologie Generala Lucrare PracticăDocument2 paginiFarmacologie Generala Lucrare PracticăAlice DouglasÎncă nu există evaluări
- Imunomodulatoare - FarmacologieDocument17 paginiImunomodulatoare - FarmacologieAlice DouglasÎncă nu există evaluări
- Chimioterapia AntiinfecțioasăDocument49 paginiChimioterapia AntiinfecțioasăAlice DouglasÎncă nu există evaluări
- Abdomenul Acut ChirurgicalDocument21 paginiAbdomenul Acut Chirurgicalbloodtype100% (7)