Documente Academic
Documente Profesional
Documente Cultură
HEMATOPOIEZA1
HEMATOPOIEZA1
Încărcat de
Voinea RazvanDrepturi de autor
Formate disponibile
Partajați acest document
Partajați sau inserați document
Vi se pare util acest document?
Este necorespunzător acest conținut?
Raportați acest documentDrepturi de autor:
Formate disponibile
HEMATOPOIEZA1
HEMATOPOIEZA1
Încărcat de
Voinea RazvanDrepturi de autor:
Formate disponibile
HEMATOPOIEZA
Hematopoieza - reprezint\ ansamblul mecanismelor care asigur\
produc]ia tuturor elementelor figurate sanguine pornind de la celula stem.
Celulele sanguine sunt in cea mai mare parte diferentiate, elemente
terminale si functionale ale liniilor celulare medulare. Aceste celule au
posibilitati foarte reduase sau absente de sinteza proteica si diviziuni
celulare (hematiile si trombocitele sunt celule anucleate). Durata de viata
este scurta: cateva ore pentru neutrofile, cateva zile pentru trombocite si
cateva saptamani (120 zile) pentru eritrocite. In acelasi timp, numarul
acestor celule circulante este crescut si mentinut relativ constant.
Hematopoieza asigura deci o productie celulara foarte importanta
din punct de vedere cantitativ : in fiecare zi se produc aproximativ 10 13
celule sanguine ceea ce corespunde la peste 2 milioane de hematii/sec.
Aceasta activitate productiva este asigurata de o mica populatie de
celule din maduva osoasa numite celule suse hematopoietice.
Hematopoieza trebuie controlata foarte strict pentru a asigura men]inerea
constanta a numarului de celule sanguine in ciuda variatiilor consumurilor
importante legate de contexte patologice (hemoragii, infectii ...). Aceasta
reglare se bazeaza pe mecanisme celulare si umorale (factori de crestere)
care pot fi stimulatori sau inhibitori ai hematopoiezei
Hematopoieza cuprinde c^teva etape importante :
- autore`nnoirea celulelor stem ;
- angajarea celulelor stem `n procesul de diferen]iere [i orientare spre o
anumit\ linie celular\, trec^nd prin etapa celulelor progenitoare ;
- proliferarea, diferen]ierea [i maturarea celulelor progenitoare
conduc^nd spre un anumit tip celular matur.
I. Dezvoltarea sistemului hematopoietic/organele hematopoietice
Sediul hematopoiezei variaza in functie de etapa evolutiva. Diferitele
localiz\ri ale hematopoiezei, `n evolu]ie, nu sunt exclusive unele fa]\ de
altele, pot exista suprapuneri `ntre ele, `n timp.
Hematopoieza are dou\ etape importante : hematopoieza primitiv\ [i
hematopoieza definitiv\.
Ia. Hematopoieza primitiv\
Debuteaz\ `n ziua 19 a vietii embrionare si dureaza pana in saptamana 8.
La inceput este mezodermica, cu formarea de insule sanguine la nivelul
sacului vitelin extra-embrionar. In insulele sanguine celulele centrale se
diferentiaza in eritrocite nucleate iar celulele periferice formeaza primele
celule endoteliale.
Ib. Hematopoieza definitiva
~ncepe `n s\pt\m^na 8. Apari]ia re]elei vasculare care conecteaz\ sacul
vitelin de embrion [i diferitele organe [i ]esuturi embrionare `ntre ele
permite trecerea celulelor stem hematopoietice din agregatele primitive,
`n circula]ia embrionar\ [i, apoi, `n orgnele hematopoietice. ~n cursul
vie]ii fetale precoce, dup\ a [asea s\pt\m^n\ de gesta]ie [i pe parcursul
celui de al doilea trimestru de sarcin\, organele hematopoietice principale
sunt : ficatul [i splina. Ulterior, organul hemetopoietic principal devine
medulara osoas\.
La na[tere, cavit\]ile medulare ale oaselor reprezint\ situsul
hematopoietic major. Toate oasele sunt implicate `n procesul
hematopoietic. ~n evolu]ie, procesul hematopoietic activ se restr^nge la
oasele scurte [i late (calota cranian\, stern, coaste, oasele bazinului,
vertebre). ~n rest, m\duva osoas\ ro[ie, hematoformatoare este `nlocuit\
cu m\duv\ gras\, galben\. La acest nivel, predomin\ adipocitele. Totu[i, la
acest nivel, ca [i `n alte organe ale sistemului reticuloendotelial, r\m^n
celule stem multipotente ca celule hematopoietice `n repaus. Aceste
celule `[i p\streaz\ capacitatea de activare astfel c\, `n situa]ii de
necesitate, sau patologice (sindroame mieloproliferative, anemii
hemolitice) s\ dea na[tere la focare hematopoietice extramedulare.
Varsta individului
Situs-ul hematopoiezei
Embrion
Sacul vitelin apoi ficatul
a-3 a-7
utero
luna in
Splina
a-4a a-5a luna in Cavitatea medulara - predominant granulocite [i
utero
plachete
a-7a luna
Cavitatea medulara - eritrocite
Na[tere
Predominant medular; splina [i ficatul in caz de
necesitate
Na[tere maturitate
Num\rul de situs-uri active in maduva osoas\
scade dar `[i men]in capacitatea hematopoietic\
Adult
m\duva osoas\ active\ la nivel craniu, coaste,
stern, vertebre, pelvis, extremit\]ile proximale
ale femurului
II. M\duva osoas\ la adult
M\duva osoas\ (MO) este localizat\ `n cavitatea a numeroase oase. De[i
este dispersat\ din punct de vedere anatomic anatomic, prezint\ o unitate
de organizare datorit\ particularit\]ilor vasculariza]iei sale [i re]elei de
celule stromale care stimuleaz\ hematopoieza.
La dult, MO activ\ reprezint\ aproximativ 50 - 60% din volumul
medular total [i se limiteaz\ la oasele craniului, clavicule, vertebre, stern,
bazin, extremitati proximale ale femurului si humerusului. ~n oasele cu
hematopoiez\ activ\, se produce o involu]ie adipoas\ mai marcat\ `n partea
central\ a osului [i se accentueaz\ cu `mb\tr^nirea.
Din punct histologic organizatoric, MO cuprinde mai multe
compartimente :
- Un cadru osos, cu celule asociate ;
- Un compartiment vascular cu anumite particularit\]i
- Parenchimul hematopoietic.
- Un micromediu sau trama conjunctivo-vasculara care cuprinde celulele
stromale [i matrice extracelulare
II.1. Cadrul osos
Este un os spongios, compus de o `ntret\iere de lamele osoase sau
trabecule care delimiteaz\ mici spatii (lojete) [i diverse celule : osteobla[ti,
osteocite, osteoclaste.
- Osteobla[tii - celule de origine mezenchimatoas\ care secret\
tropocolagen (precursor de colagen ce va forma trama osteoida),
glicoproteine, mucopolizaharide [i factori de crestere : GM-CSF, M-CSF,
IL1, TGF.
- Osteocite derivate din osteoblaste. Sunt con]inute `n cavit\]i numite
osteoplaste ; particip\ la nutri]ia [i activitatea osoas\.
- Osteoclaste celule de origine monocitar\macrofagic\ cu rol `n
osteoliz\ rezorb matricea mineral\ osoas\ prin secre]ia de acizi
organici care solubilizeaz\ s\rurile de calciu. Sunt stimulate de
parathormon [i de factorul activator al osteoclastelor (OAF) `n contexte
patologice.
Viata osului este ciclic\ cu edific\ri [i rezorb]ii succesive (reinnoirea
total\ a structurii osoase se realizeaz\ `n aproximativ 20 ani). Osul este
format initial sub o forma nemineralizata (os osteoid) apoi mineralizarea
survine in mod secundar.
II. 2. Vascularizatia osoasa
Are un rol functional : nutritia, oxigenarea, productia de factori de crestere,
si eliberare de celule mature.
Sinusurile venoase medulare sunt formate dintr-un monostrat
continuu de celule endoteliale ale c\ror jonc]iuni sunt realizate printr-o
juxtapozi]ie celular\ cu o discret\ suprapunere a marginilor citoplasmatice.
Pe fa]a extraluminal\, celulele endoteliale sunt acoperite de un strat
discontimuu format de celule reticulare adventiceale (fibrobla[ti sau
miofibrobla[ti care prezint\ o serie de prelungiri ce formeaz\ o re]ea extins\
`n spa]iul medular extravascular). Migrarea celulelor hematopoietice intra
[i extravascular, la nivel medular se realizeaz\ la nivelul sinusurilor
venoase. Un rol esen]ial `l joac\ celulele endoteliale care regleaz\ trecerea
intravascular\ selectiv\ a celulelor sanguine mature, c^t [i trecerea
intramedular\ a progenitorilor hematopoietici, `n special dup\ grefa
medular\.
II.3. Parenchimul hematopoietic :
Din punct de vedere structural se caracterizeaz\ prin juxtapozi]ia de
diverse celule mieloide care se reg\sesc `n mici regiuni definite de c\tre
lamelele osoase (lojete medulare). Acestea con]in o tram\ (fibre ale
matricei extra-celulare [i fibrobla[ti), adipocite, celule endoteliale, capilare
sinusoide si ansamblul de elemente imature si diferen]iate ale
hematopoiezei. Regiunile endostale juxtatrabeculare sunt mai bogate in
precursori hematopoietici ; compartimentele de maturatie sunt mai
`ndepartate. ~n patologie, `n sindroamele mielodisaplazice, se constat\
focare de celule imature la distan]\ de endost [i de capilarele sinusoide
(ALIP).
Din punct de vedere functional, parenchimul hematopoietic se
organizeaza in mai multe compartimente ierarhizate :
celulele stem hematopoietice
celulele progenitoare
celulele precursoare
a) Celulele stem
Produc]ia permanent\ a tuturor celulelor mature din s^nge este asigurat\
de o popula]ie celular\ medular\, destul de restr^ns\ numeric, capabil\ de
autore`nnoire, de proliferare intens\ [i diferen]iere celulele stem
hematopoietice totipotente. Capacitatea de autore`nnoire asigur\
men]inerea unei rezerve constante de celule stem. Capacitatea
proliferativ\ crescut\ asigur\ un num\r sporit de celule descendente. Totu[i,
nu trebuie uitat c\ num\rul de diviziuni succesive la care particip\ o celul\
stem este limitat. Un studiu experimental a ar\tat c\ o celul\ stem poate da
na[tere la peste 500.000 celule diferen]iate, dup\ doar 19 diviziuni
succesive.
Aceste celule nu pot fi identificate morfologic, cel pu]in cu tehnicile
actuale. Se crede c\ sunt celule mononucleate, cu un diametru de 6
microni [i un raport nucleo-citoplasmatic foarte ridicat. Marea majoritate
sunt `n repaus celular (faza G 0 a ciclului celular). Recunoa[terea celulei se
face pe baza analizei fenotipului imunologic (markerii de suprafa]\
exprima]i pe membrana celular\) : CD34+, CD38-, DR-, CD45RA-, c-kit+,
CD19-. Recent au fost descoperite dou\ antigene noi, exprimate pe
membrana acestor celule : HCA (molecul\ de adeziune) [i HCC-1.
Celulele hematopoietice pot trece `n s^ngele periferic, fie `n etapa
de recuperare a unei neutropenii postchimioterapie, fie sub efectul
stimulator al factorilor de cre[tere hematopoietici. Se pare c\ aceste celule
hematopoietice sanguine ar fi mai imature dec^t celulele hematopoietice
medulare prelevate `n faz\ stabil\ (`nafara situa]iilor men]ionate mai sus).
Celula stem totipotent\, aflat\ `n repaus (faza G0), sub ac]iunea unor
factori stimulan]i, va intra `n ciclul celular conduc^nd la diviziunea
celular\. Rezultatul diviziunii vor fi celule stem identice [i/sau celule
angajate. Exist\ opinii diferite, potrivit c\rora diviziunile celulare
conduc^nd la cele dou\ tipuri de celule fiice sunt independente, `n func]ie
de stimul. Al]i autori sunt de p\rere c\ diviziune celulei stem totipotente
este asimetric\ conduc^nd la dou\ celule fiice diferite : una identic\ cu ea
`ns\[i (autore`nnoirea cu men]inerea rezervei) [i una angajat\ `n
procesul de diferen]iere, spre anumite linii celulare. Celula angajat\
rezultat\ `n prima genera]ie are poten]ialul de a da descenden]i
apar]in^nd mai multor linii celul\ stem pluripotent\ sau multipotent\
(mieloid\ sau limfoid\). ~n etapa urm\toare procesul de diferen]iere
continu\, cu restric]ia la o singur\ linie celular\ - celula stem unipotent\.
Deci, celula stem va suferi un proces de diferen]iere [i proliferare.
Proliferarea va asigura un num\r crescut de celule descendente.
Diferen]ierea sau angajarea va determina orientarea treptat\ spre o
anumit\ linie celular\ sanguin\. Acest proces ireversibil va determina
pierderea capacit\]ii de re`nnoire [i limitarea posibilit\]ilor de evolu]ie spre
diferitele elemente mature.
b) Celulele progenitoare
Odat\ procesul de angajare ini]iat, celulele intr\ `n compartimentul
celulelor progenitoare, compartiment intermediar `ntre cel al celulelor
stem [i al celulelor precursoare.
Celulele progenitoare au o ierarhie proprie, de la cele mai precoce,
cu o gam\ mai larg\ de posibilit\]i evolutive, spre cele tardive, mature cu o
singur\ posibilitate evolutiv\. Se pare c\ procesul de angajare are o
ordine bine definit\ a etapelor. O prim\ etap\ este angajarea pe linia
mieloid\ [i respectiv limfoid\, d^nd celule progenitoare pluripotente (CFUGEMM [i CFU-L).
Ulterior procesul de diferen]iere progreseaz\ cu restr^ngerea paletei
evolutive. Fiecare celul\ progenitoare va da na[tere la un repertoriu bine
definit [i limitat de celule mature. Fiecare celul\ progenitoare `[i p\streaz\
capacitatea proliferativ\ [i va suferi multiple diviziuni la fiecare etap\, p^n\
la intrarea `n compartimentul de celule precursoare. ~n paralel cu
procesul de multiplicare se continu\ [i cel de diferen]iere, dar capacitatea
de diferen]iere se restr^nge de la o etap\ la alta.
Aceste celule progenitoare cuprind : CFU-E (colony forming units
erytrocyte), CFU-G (granulocyte), CFU-M (monocyte/macrophage), CFU-MK
(megakariocyte), CFU-preB (B lymphocytes), CFU-DL (dendritic/langerhans
cell). ~n anumite linii celulare, cum este cazul celei eritrocitare, pot exista
mai multe etape de matura]ie. Toate aceste celule nu pot fi identificate
morfologic la examenul microcscopic, dar pot fi identificate prin
capacitatea lor de a forma colonii celulare.
c) Celule precursoare
Celulele precursoare reprezint\ stadiile finale ale hematopoiezei. Celulele
sunt destul de diferen]iate pentru a fi identificate morfologic la examenul
microscopic pe frotiuri medulare. Capacitatea de proliferare s-a redus la
maximum `nc^t celulele mai pot suferi, `n medie, patru diviziuni celulare.
~n paralel, celulele sufer\ modific\ri asociate cu ultimile etape matura]ie
care conduc la elementele figurate mature. Modific\rile se refer\ la
dimensiuni (dimensiunile celulei scad pe m\sura diviziunilor ultime)
aspectul nuclear (nucleul `[i mic[oreaz\ dimensiunile, cromatina se
condenseaz\ `n gr\mezi, nucleolii, prezen]i la `nceput, dispar) [i aspectul
citoplasmatic (ini]ial citoplasma este intens bazofil\ datorit\ `nc\rc\rii cu
ribozomi [i sintezei proteice intense, ulterior tinctorialitatea vireaz\ treptat
spre acidofilie, [i se `ncarc\, eventual, cu granula]ii). Acestea vor p\r\si
m\duva trec^nd `n circula]ie pentru a compensa pierderile cotidiene.
II.4. Stroma medular\ (Micromediul medular)
Hematopoieza la adult are loc `n medulara oaselor late [i scurte. Biopsiile
osoase au ar\tat c\ celulele hematopoietice sunt localizate `n
compartimente situate `ntre trabeculele osoase. La acest nivel, `nafar\ de
aceste celule hematopoietice [i structurile vasculare, mai pot fi puse `n
eviden]\ elemente celulare nonhematopoietice [i structuri acelulare. Toate
acestea, `n ansamblu, alc\tuiesc stroma medular\ sau micromediul
medular, concept important `n `n]elegerea mecanismului hematopoiezei.
Stroma este alc\tuit\ din :
elemente celulare : miofibrobla[ti, adipocite, celule endoteliale,
limfocite, plasmocite, macrofage ;
matricea extracelular\ : proteine fibroase (colagen I, III, IV, V, VI,
fibronectina),
glicoproteine
(laminina,
hemonectina,
nascina,
trombospondina) [i proteoglicani.
Stroma sau micromediul medular joac\ un dublu rol esen]ial `n
procesul de hematopoiez\:
asigur\ fenomenul de homing $ (de localizare) la nivel medular al
celulelor stem hematopoietice [i deci al hematopoiezei. Astfel, pe de o
parte, se asigur\ prin anumite semnale atragerea celulelor stem cu
colonizarea spa]iilor medulare osoase. Colonizare care are loc fie `n
procesul ontogenetic, fie `n cazul celulelor stem toti- sau multipotente
recirculate prin s^ngele periferic, fie `n caz de gref\ medular\. Pe de alt\
parte, se asigur\ fixarea celulelor hematopoietice la nivel medular,
limit^nd trecerea celulelor `n s^ngele periferic doar pentru elementele
mature. Acest fenomen implic\ un proces de adeziune a celulelor
hematopoietice la celulele [i matricea stromal\. Rezult\ o interac]iune
direct\ celul\-celul\. Celulele hematopoietice [i cele stromale exprim\ la
nivel membranar molecule de adeziune car permit fixarea intercelular\
[i ancorarea la matrice.
Celulele stromale produc [i elibereaz\ numero[i factori de cre[tere [i
inhibitori care moduleaz\ procesul de hematopoiez\.
$ Celulele hematopoietice progenitoare posed\ la nivel membranar receptori care se leag\
`n mod specific la situsuri particulare de pe macromoleculele matricei. Astfel, matricea
extracelular\ asigur\ o re]ea structural\ la care se fixeaz\ progenitorii, particip^nd la
fenomenul de localizare (homing). Odat\ cu apropierea de fazele terminale ale maturiz\rii,
celulele hematopoietice `[i pierd aceste molecule membranare care interac]ioneaz\ cu
matricea, permi]^nd migrarea extramedular\. Proteinogliganii intens sulfata]i ca heparan
sulfatul [i condroitin sulfatul contribuie la adeziunea `ntre strom\ [i celule hematopoietice.
Proteinoglicanii pot ac]iona prin concentrarea factorilor de cre[tere solubili, ajut^nd
hematopoieza.
IV. Factorii de cre[tere hematopoietic\
Factorii de cre[tere hematopoietic\ reprezint\ un grup heterogen de
citokine
care
stimuleaz\
celulele
progenitoare
ale
sistemului
hematopoietic. Drept consecin]\ se ini]iaz\ procesul de diferen]iere,
proliferare [i maturare al acestor celule. Marea lor majoritate sunt
glicoproteine. Sunt sintetizate [i elaborate (cu excep]ia eritropoietinei) de
c\tre celulele stromei medulare dar [i din alte organe (celulele endoteliale,
fibrobla[ti, monocite/macrofage, limfocite). Factorii de cre[tere se leag\
specific de receptori corespunz\tori de pe membrana celulelor
hematopoietice. Ei joac\ un rol esen]ial `n reglarea hematopoiezei, at^t `n
perioadele de s\n\tate, c^t [i pe timp de boal\.
Grupul factorilor de cre[tere este extrem de heterogen at^t din
punct de vedere al structurii chimice primare, dar [i din punct de vedere
func]ional. Sunt factori de cre[tere capabili s\ ac]ioneze pe mai multe linii
celulare (Il-3, GM-CSF), dup\ cum exist\ factori cu specificitate pe anumite
linii celulare (EPO, G-CSF, M-CSF). Exist\ factori care influen]eaz\ toate
etapele devenirii celulelor hematopoietice, dup\ cum exist\ factori de
cre[tere care ac]ioneaz\ punctual, fie `n stadiile precoce asigur^nd
recrutarea celulelor aflate `n repaos [i introducerea `n ciclul celular cu
ini]ierea angaj\rii celulare [i prolifer\rii, fie ac]ioneaz\ numai `n stadiile mai
tardive fiind necesare diferen]ierii [i matur\rii celulare. Astfel procesul de
hematopoiez\ necesit\ colaborarea tuturor acestor factori pentru ini]ierea [i
finalizarea sa.
Factorii de cre[tere `[i pot exercita ac]iunea reglatoare asupra
hematopoiezei at^t `n mod endocrin (ac]iunea la distan]\, pe cale
sanguin\), `n mod paracrin (prin contiguitate, fie prin secre]ie, fie prin
contact membranar), dar [i `n mod autocrin (fie prin secre]ie, celula
posed^nd [i receptorul membranar corespunz\tor, fie direct intracelular).
Mul]i dintre factorii de cre[tere au fost identifica]i, ca [i gena lor
codant\. Gena a fost clonat\ [i astfel s-a putut trece la produc]ia lor pe
scar\ larg\ prin inginerie genetic\. Acest lucru a permis stuierea lor [i
studiul in vitro al procesului de hematopoiez\ `n scopul amelior\rii
`n]elegerii lui. ~n acela[i timp a permis utilizarea lor `n terapeutic\ :
colectarea celulelor stem pentru gref\, diminuarea duratei [i profunzimii
aplaziilor post-chimioterapie, post-gref\
a) Controlul pozitiv al hematopoiezei
~n ansamblu, hematopoieza este controlat\ `n mod pozitiv de :
factori cu ac]iune sinergic\, care ac]ioneaz\ asupra celulelor stem
totipotente (capabile de autore`nnoire) IL-3, IL-1, IL-6 ;
factori
care ac]ioneaz\ asupra celulelor pluripotente [i asupra
progenitorilor cei mai diferen]ia]i IL-3, IL-4, GM-CSF ;
factori care ac]ioneaz\ asupra etapelor tardive ale diferen]ierii [i
matur\rii, av^nd specificitate pentru anumite linii celulare G-CSF, M-CSF,
Epo, MK-CSF sau Tpo, IL-5.
Factorii de cre[tere se fixeaz\ pe receptori specifici, exprima]i pe
suprafa]a membranei celulare. Receptorii identifica]i p^n\ `n prezent
apar]in de dou\ mari familii de receptori : receptori cu activitate tirozinkinaz\ [i superfamilia receptorilor citokinelor. Legarea de receptori a
factorilor de cre[tere, determin\ declan[area unei cascade de reac]ii
chimice [i interven]ia unor mesageri intracelulari care vor conduce la
realizarea r\spunsului corespunz\tor. Fiecare factor de cre[tere `[i are
propriul receptor, diferit de al celorlal]i. ~n acela[i timp, o celul\
hematopoietic\ poate exprima receptori pentru mai mul]i factori de
cre[tere.
Rolul capital al factorilor de cre[tere asupra celulelor hematopoietice
este acela de a asigura proliferarea, diferen]ierea [i supravie]uirea
celulelor ]int\. 1) Factorii de cre[tere asigur\ antrenarea celulelor aflate `n
repaos [i intrarea lor `n ciclul celular proliferant, ac]ion^nd mai ales `n
timpul stadiului G1. 2) Factorii de cre[tere pot activa factori de transcrip]ie
ai unor gene cu declan[area sintezei proteinelor codate [i, astfel, ini]iaz\
procesul de angajare [i apoi diferen]iere pe diferitele linii celulare. 3) Un
element important de subliniat este rolul de asigurare a suprvie]uirii
celulare pentru c\, absen]a factorului de cre[tere nu antreneaz\ numai
oprirea ciclului celular ci [i moartea celular\. Astfel, factorul de cre[tere
previne declan[area apoptozei, probabil prin reglarea expresiei genei bcl-2,
implicat\ `n acest proces.
b) Controlul negativ (inhibitor) al hematopoiezei
Ca orice proces din organism, [i hematopoieza este controlat\. Controlul
exercitat este at^t pozitiv (stimulant) c^t [i negativ (inhibitor). Echilibrul
fin `ntre cele dou\ sisteme de control asigur\, `n condi]ii fiziologice, o
hematopoiez\ normal\ (cantitativ [i calitativ). Orice dereglare a unuia din
mecanisme poate antrena hematopoieza `n una din direc]ii de produc]ie
excesiv\ sau deficitar\.
Sistemul de control negativ este mai pu]in bine cunoscut dar
progrese importante s-au realizat `n ultimii ani. Cunoa[terea acestui
sistem prezint\ o serie de interese : a) o mai bun\ cunoa[tere a controlului
hematopoiezei normale ; b) explicarea modului de implicare a inhibi]iei
hematopoiezei normale `n cursul leucemiilor ; c) posibila utilizare a
factorilor inhibitori pentru scoaterea din ciclul celular a celulelor stem `n
timpul tratamentului citostatic [i, astfel, protejarea capitalului
hematopoietic.
Ca [i factorii stimulan]i, factorii inhibitori ac]ioneaz\ la mai multe
nivele ale prolifer\rii [i diferen]ierii hematopoietice. Ei nu au specificitate
de linie celular\.
Factori de control negativ :
Prostaglandinele E
Interferonii
Tumor Necrosis Factor alfa (TNF alfa)
Transforming growth factor beta (TGF beta)
Macrophage inflammatory protein (MIP1)
Isoferitinele acide
Lactoferina
Tetrapeptidul acetil-N-Ser-Asp-Lys-Pro (AcSDKP, Seraspenid)
Pentapeptid pGlu-Glu-Asp-Cys-Lys (pEEDCK)
Factorul plachetar 4 (PF4)
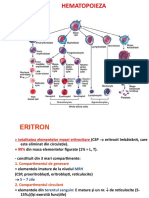
A. ERITROPOIEZA
Eritrocitele la adultul normal provin din celulele su[e (stem) din m\duva
osoas\ hematopoietic\ care, dup\ ce s-au diferen]iat, sufer\ un proces de
matura]ie ce dureaz\ n jur de [ase zile. ntregul proces prin care
eritrocitele sunt produse la nivelul m\duvei osoase se nume[te eritropoiez\.
Eritropoieza este un proces controlat care asigur\ o produc]ie continu\ ce
nlocuie[te pierderile fiziologice men]in^nd constant\ masa eritrocitar\
total\ din organism. Eritropoieza implic\ un num\r [i o varietate mare de
celule n diferite etape de matura]ie ncep^nd de la prima celul\ stem
angajat\ spre linia eritrocitar\ p^n\ la eritrocitul matur. Aceast\ ntreag\
mas\ de celule eritroide a fost denumit\ eritron. Acest concept prive[te
masa eritroid\ ca o unitate tisular\ func]ional\.
Eritropoieza cuprinde mai multe etape. Celula stem multipotent\ sub
ac]iunea unor stimuli se angajeaz\ spre linia eritroid\. Celulele rezultate vor
suferi ini]ial un proces de proliferare [i diferen]iere n etape iar ulterior un
proces de matura]ie. Celulele cele mai precoce ale liniei ro[ii formeaz\
compartimentul celulelor progenitoare cuprinznd BFU-E (burst-forming
unit-erythroid) [i CFU-E (colony-forming unit-erythroid). Aceste celule nu
pot fi recunoscute din punct de vedere morfologic dar pot fi identificate
func]ional prin capacitatea lor de a forma in vitro colonii eritroblastice.
Procesul se continu\ cu compartimentul celulelor precursoare ce cuprinde
n ordine maturativ\ : proeritroblastul, eritroblastul bazofil, eritroblastul
policromatofil, eritroblastul acidofil, pentru a ajunge la reticulocit [i, n
final, eritrocitul matur.
Celulele progenitoare :
Burst Forming Unit Erythroid (BFU-E) apare morfologic ca un blast
foarte imatur, u[or ovalar, cu citoplasma moderat bazofil\, uneori cu
pseudopode fine, nucleul mare cu cromatina foarte fin\ [i nucleoli mari.
Colony Forming Unit Erythroid (CFU-E) apare morfologic ca o celul\
imatur\, cu nucleu mare cu cromatina nuclear\ fin\, nucleoli mari,
eviden]i, o zon\ perinuclear\ clar\ [i citoplasm\ bazofil\ cu pseudopode.
Celulele precursoare :
Proeritroblastul - reprezint\ cea mai precoce celul\ a seriei ro[ii posibil
de identificat cu ajutorul tehnicilor actuale. Este o celul\ mare, rotund\
sau ovalar\, cu diametrul de 20-25 m. Nucleul este rotund sau oval cu
cromatina fin granular\ [i unul sau doi nucleoli. Ocup\ aproximativ 80%
din celu\. Citoplasma are o culoare albastru nchis. Proeritroblastul
sufer\ mitoz\ d^nd na[tere la dou\ celule fiice ce se matureaz\ n
eritrobla[ti bazofili.
Eritroblastul bazofil - talia se mic[oreaz\ : 16-18 m. Cromatina nuclear\
ncepe s\ se condenseze n gr\mezi dispuse predominant la periferia
nucleului. Nucleolii sunt absen]i. Citoplasma are o culoare albastru
nchis. Are loc o sintez\ crescut\ a hemoglobinei n citoplasm\ dar
cantitatea crescut\ de ARN o mascheaz\.
Eritroblastul policromatofil - raportul nucleo/citoplasmatic continu\ s\
scad\, ca [i talia celulei, de numai 9-12 m. Nucleul s-a mic[orat, cu
cromatina condensat\. Sinteza de hemoglobin\ este crescut\ cu invazia
treptat\ a citoplasmei nc^t colora]ia este variabil\, n general albastrugri. Acesta este ultimul stadiu c^nd se mai produc mitoze, duc^nd la
urm\toarea etap\.
Eritroblastul acidofil sau ortocromatofil - Nucleul complet picnotic,
incapabil de sintez\ a ADN este expulzat din celul\ (kariorexis).
Citoplasma este aproape complet ocupat\ de hemoglobin\, devenind
roz-portocalie.
Reticulocitul - reprezint\ stadiul premerg\tor eritrocitului matur.
Citoplasma seam\n\ cu cea a eritroblastului acidofil. R\m^n cantit\]i
variabile de ARN d^nd culoarea alb\struie. Reticulocitele pot p\r\si
m\duva [i trec n circula]ie. Ele sunt eviden]iabile pe frotiul din s^ngele
periferic prin colora]ii speciale (ex. albastru de metilen sau albastru de
cresyl) care pun n eviden]\ o re]ea reticulo-filamentoas\, prin
precipitarea poliribozomilor. n aceast\ etap\ se pierde [i restul de
organite celulare ajung^nd la stadiul terminal.
Eritrocitul - celula matur\ circulant\. Are aspect de disc biconcav cu
diametrul de 7-8 m, reprezent^nd forma cea mai avantajoas\ din
punct de vedere al raportului suprafa]\/volum, ce asigur\ o nc\rcare
optim\ cu hemoglobin\. Pe frotiu apare ca o celul\ rotund\, f\r\ nucleu, cu
citoplasma ro[ie-roz, mai palid\ central [i mai intens\ periferic. Durata
medie de via]\ este de 120 zile. Func]ia principal\ este de transport al
oxigenului de la pl\m^ni la ]esuturi. De aceea interiorul celulei con]ine
90% hemoglobin\ [i 10% ap\.
Eritropoieza, ca ntreaga hematopoiez\, se afl\ sub controlul factorilor
de cre[tere, n principal sub controlul eritropoietinei cu specificitate
aproape exclusiv\ pentru linia ro[ie. Eritropoietina este un hormon
glicoproteic produs n principal la nivel renal, pare-se de celulele
intersti]iale din apropierea tubului contort distal. Produc]ia hepatic\,
esen]ial\ la f\t, reprezint\ doar 10-15% la adult. Sinteza eritropoietinei este
sub controlul presiunii O2 la nivel tisular.
B. GRANULOCITELE NEUTROFILE
Granulocitele din s^nge sunt celule care, a[a cum arat\ [i numele, con]in
`n citoplasm\ numeroase granula]ii. ~n func]ie de afinitatea tinctorial\, se
disting trei tipuri de granula]ii, care dau [i numele celor trei subpopula]ii
de granulocite : neutrofile, eozinofile [i bazofile. Granulocitele neutrofile
sunt cele mai numeroase.
Granulocitele se formeaz\ `n m\duva osoas\ hematopoietic\. Ele iau
na[tere, ca [i seria eritrocitar\ [i megacariocitar\, dintr-o celul\ su[\
pluripotent\, care se diferen]iaz\ spre progenitorul specific liniei. Procesul
de matura]ie la nivel medular dureaza aproximativ 7 zile [i cuprinde `n jur
de 4-5 mitoze. Procesul de diferen]iere [i maturare se traduce morfologic
prin schimb\ri de form\ ale nucleului [i o maturare progresiv\ a citoplasmei,
cu pierderea bazofiliei [i apari]ia de granula]ii specifice.
La nivel medular, `n cadrul procesului de maturare, pot fi
diferen]iate mai multe etape pe linia celulelor precursoare : mieloblast,
promielocit, mielocit, metamielocit, granulocit nesegmentat [i granulocitul
matur sau polimorfonuclearul (PN).
Neutrofilele constituie 40-70% din totalitatea globulelor din s^nge.
Linia lor de matura]ie constituie popula]ia majoritar\ din maduv\. Raportul
granulocite/eritrobla[ti este de aproximativ 3/1.
1. Mieloblastul.
Constituie celula cap de serie, recognoscibil\ morfologic `n microscopia
optic\, a seriei precursoare granulocitare. Este o celul\ prezent\ `n
procentaj sc\zut `n m\duva normal\ (0,5-2%). Are un diametru de 15-20
microni (etalat\). Este oval\, uneori rotunjit\. Nucleul are cromatina
omogen\, fin dispersat\, cu nucleoli (2-5) bine delimita]i. Citoplasma este
albastr\, mai `nchis\ la periferie, cu rare granula]ii azurofile, de talie mic\.
Num\rul granula]iilor cre[te cu maturarea.
Specializarea fiecarei linii celulare apare precoce, `nc\ din aceast\
etap\, dar nu poate fi pus\ `n eviden]\, `n microscopia optic\, dec^t din
stadiul de mielocit (granula]iile specifice).
2. Promielocitul.
Este o celul\ mai voluminoas\ dec^t mieloblastul. Este prezent\ `n
propor]ie de 2-8% din popula]ia medular\. Nucleul este voluminos, adesea
excentric, discret anco[at, cu cromatina ce `ncepe s\ se condenseze, iar
nucleolii sunt uneori vizibili. Citoplasma este abundent\, mai pu]in bazofil\
ca la mieloblast [i bogat\ `n granula]ii. ~n vecin\tatea nucleului este o
zona mai s\rac\ `n granula]ii, unde este situat aparatul Golgi.
3. Mielocitul.
10
Este o celul\ de 15 microni, cu un nucleu rotunjit, cu cromatina condensat\
`n gr\mezi. Citoplasma este acidofil\, bogat\ `n granula]ii neutrofile, de
culoare maronie deschis. Granula]iile azurofile, de[i `nc\ prezente, sunt
`nvelite `n mucopolizaharide cu pierderea capacit\]ilor tinctoriale `nc^t nu
mai sunt vizibile.
4. Metamielocitul.
Celula se caracterizeaz\ prin alungirea [i `ncurbarea nucleului, cu
cromatina condensat\ `n blocuri bine delimitate. Citoplasma este acidofil\
cu aspect similar cu a mielocitului. Urmeaz\ o etap\ intermediar\ de
neutrofil nesegmentat (band neutrophil - din literatura anglo-saxon\) `n
care nucleul alungit este total `ncurbat, lu^nd aspectul de potcoav\.
5. Polinuclearul neutrofil.
Aspectul pe frotiu este al unei celule rotunde cu diametrul de 12-14
microni. Nucleul este format segmente (blocuri) cromatiniene (2-5) reunite
prin filamente fine. ~n s^ngele periferic, 50% din neutrofilele mature au 3
lobi, [i sub 5% au 5 lobi sau peste. Citoplasma este roz, plin\ cu
numeroase granula]ii neutrofile. 70-80% dintre granula]ii sunt neutrofile,
iar restul sunt azurofile.
C. SERIA MONOCITO - MACROFAGICA
Studiul modern al fagocitelor la mamifere a fost `nceput secolul trecut de
c\tre Metchnikoff. Studiul ontogeniei, cineticii, [i func]iei celulelor
fagocitante la animale a determinat crearea conceptului de sistem
monocitic-macrofagic sau sistemul fagocitic mononuclear. Acest sistem
cuprinde monobla[tii [i promonocitele din m\duva osoas\, monocitele
sanguine, [i macrofagele libere [i fixate `n ]esuturi.
Acest concept de sistem fagocitic mononuclear exclude
polinuclearele, ca celule fagocitante, [i `nlocuieste conceptul mai vechi al
lui Aschoff din 1924, de sistem reticulo-endotelial care cuprindea al\turi de
macrofage, celulele endoteliale, celulele reticulare, celulele dendritice din
centrii germinativi ai foliculilor ganglionari. Aceste celule joac\ un rol
complementar `n activitatea fagocitar\ dar au origini diferite fa]\ de
sistemul monocitic-macrofagic.
Produc]ie
Sistemul fagocitic mononuclear cuprinde monoblastul, promonocitul,
monocitul si macrofagul. ~[i are originea, ca [i restul elementelor figurate
sanguine, la nivelul m\duvei osoase hematoformatoare.
Produc]ia debuteaz\ prin progenitori foarte nediferentiati =
CFU-GEMM
(Colony
Forming
Unit
Granulocitar,
Eritroblastic,
Megacariocitar si Monocitar) care, sub influen]a diverselor citokine (SCF,
FLT3-L, GM-CSF, IL-3) vor da na[tere la progenitori diferen]ia]i.
Acesti progenitori mieloizi mai diferentiati (CFU-GM)
(Granulocitari [i monocitari), vor prolifera `n prezen]a IL-3 [i GM-CSF
(Granulocyte Monocyte - CSF). Apoi, prezenta suplimentara de G-CSF
stimuleaz\ orientarea spre CFU-G (care vor da na[tere la linia granulocitar\
neutrofil\), `n timp ce prezen]a suplimentar\ de M-CSF determin\
diferen]ierea spre progenitorii CFU-M apoi spre precursorii monocitari.
11
Timpul de diferentiere al CFU-GEMM in monocite medulare = aprox
4-5 zile.
Macrofagul
constituie
elementul
central
i`n
reglarea
monocitopoiezei. Macrofagul activat de diver[i stimuli (imuni, endotoxina)
este capabil s\ produc\ M-CSF [i GM-CSF ce vor stimula monocitopoieza.
~n acela[i timp macrofagul produce citokine (TNF, IL-1) care vor stimula
celulele endoteliale [i mezenchimale s\ produca M-CSF [i GM-CSF.
~n acela[i timp GM-CSF interac]ioneaz\ cu macrofagul matur [i `l
activeaz\ cresc^ndu-i poten]ialul citotoxic. Limfocitul T activat secret\ GMCSF [i IL-3 care vor ac]iona local [i vor regla proliferarea [i activarea
macrofagelor `n focar.
Morfologie
CFU-GM constituie celula progenitoare a seriei monocitare. Monoblastul [i
promonocitul sunt celulele precursoare ale seriei.
1. Monoblastul normal este o celul\ rar\ la nivelul m\duvei osoase normale.
Microscopia optic\ aproape nu poate face diferen]ierea cu mieloblastul.
Monoblastul devine evident `n situatii patologice, cum este cazul
leucemiei acute monoblastice (LAM5). ~n general, este o celul\ cu aspect
blastic, celul\ t^n\r\ cu nucleu mare cu cromatin\ fin granular\ [i uniform
dispersat\, [i cu nucleoli prezen]i.
2. Promonocitul reprezint\ celula intermediar\ `ntre monoblast [i monocit.
Nucleul este puternic anco[at, neregulat [i prezint\ filamente `n m\nunchi
sau dispersate `n citoplasm\.
Pentru o mai corect\ diferen]iere este util\ folosirea reac]iilor
citochimice (valabile [i pentru monocit). Celulele contin mieloperoxidaza
ca [i linia granulocitara dar `n cantitate mult mai mic\. Reac]iile
citochimice pot pune in eviden]\ o reac]ie fosfatazic\ acid\, si mai ales
esterazic\. Aceasta din urm\ este foarte util\ fiind cu specificitate pentru
linia monocitar\ deoarece este inhibat\ de florura de sodiu, fenomen care
nu se petrece `n cazul esterazelor din linia granulocitar\.
3. Monocitul este cea mai mare celula sanguin\, are un diametru de 12-15
microni. Nucleul ocup\ aproximativ jum\tate din aria celular\ [i este
pozi]ionat, de obicei, excentric. Nucleul este adesea reniform sau puternic
anco[at, neregulat, mai rar rotund dar cu contur neregulat. Cromatina este
structurat\ `n gr\mezi situate pe fa]a intern\ a membranei nucleare, unite
prin benzi fine cromatiniene. Citoplasma are o culoare cenu[ie, grialbastruie [i con]ine fine granula]ii roz, care, uneori, pot fi numeroase
`nc^t dau citoplasmei o tent\ roz. ~n citoplasm\ pot fi prezente vacuole `n
num\r variabil.
Monocitele p\r\sesc m\duva la scurt timp dupa ultima diviziune
celular\, ceea ce explic\ faptul c\, spre deosebire de granulocite, nu exist\ o
rezerv\ medular\ de monocite. Ele trec `n circuitul sanguin. La adultul
normal monocitele reprezint\ 1-6% din popula]ia leucocitar\.
Monocitul medular r\m^ne `n maduv\ o zi dupa care migreaz\ in
s^ngele periferic unde circula 1-4 zile `nainte de a ie[i din capilare (prin
diapedeza activ\) pentru a se localiza in tesuturile din organism. Se
transforma in histiocit cu functie macrofagica si imuna. Durata de viata
este de la 3 luni la 3 ani.
12
Spre deosebire de polinuclearele neutrofile monocitele si histiocitele
sunt inca capabile sa prolifereze (chiar daca slab), dar mai ales sa
sintetizeze numeroase componente (citokine, altele..).
Potrivit localizarii lor tisulare aspectul difera [i au denumiri diferite:
Ficat = celule Kpffer
Maduva osoasa = macrofage medulare
Rinichi = celule mezangiale intraglomerulare
Creier = celulele microgliei
Seroase = macrofagele seroaselor
Plaman = macrofage alveolare
Splina = macrofage sinusale
Ganglioni = macrofage sinusale (in sinusuri), celule interdigitate
(regiunile T), celule foliculaire dendritice (centrii germinativi)
Os = osteoclaste multinucleate
Func]ii `ndeplinite :
- fagocitoza
- initierea raspunsului imun prin calitatea de celula prezentatoare de
antigene
- raspunsul inflanmator
- coagulare
D. LIMFOPOEZA
Limfoctele reprezint\ o popula]ie heterogen\ de celule care difer\ mult
`ntre ele din punct de vedere al originii, c\ii de maturare, durat\ de via]\,
teritoriile preferen]iale de populare `n organele limfoide, markeri de
suprafa]\, func]ie. Diferite aspecte morfologice ale acestor celule ca :
m\rime, granula]ii, raportul nucleo-citoplasmatic prezint\ diferen]e `ntre o
subpopula]ie limfocitat\ [i alta. Totu[i, diferen]ele sunt greu de sesizat
pentru a diferen]ia `ntre ele aceste subpopula]ii doar pe criterii
morfologice.
Cea mai important\ metod\ de diferen]iere a subpopula]iilor
limfocitare `ntre ele se bazeaz\ pe identificarea unor structuri proteice,
dintre care unele sunt glicoproteine, exprimate la nivelul membranei
celulare. Acestea constituie markeri celulari de suprafa]\ [i au fost
denumite antigene cluster of differentiation (CD). Aceste antigene
pot fi prezente pe mai multe linii celulare, unele pot avea specificitate
pentru o anumit\ linie celular\, iar altele pot avea specificitate pentru o
anumit\ etap\ din evolu]ia (maturarea) liniei celulare respective.
Identificarea lor, cu ajutorul anticorpilor monoclonali, permite identificarea
liniei [i etapei evolutive a celulei respective. Faptul este cu at^t mai
important pentru identificarea originii celulare a unei celule maligne.
~n ceea ce prive[te markerii limfocitari, au fost clasifica]i `n trei
tipuri :
- markeri de diferen]iere B sau T - cu specificitate pentru fiecare linie
limfocitar\ `n parte ;
- markeri de matura]ie structuri care apar `n anumite etape ale
devenirii celulei respective, put^nd dispare la etapele urm\toare ;
13
markeri de activare antigene care apar `n cursul ciclului celular
`ndic^nd o proliferare limfocitar\.
Limfocitele, ca [i restul celulelor sanguine, deriv\ din celula stem
hematopoietic\ totipotent\, prin intermediul unei celule progenitoare
primitive, angajate spre linia limfoid\. Diferen]ierea `n acest sens depinde
de o serie de factori celulari [i umorali (micromediul) ca [i de alte celule
sanguine specializate provenite din celula stem.
Dup\ angajarea pe linia linia limfoid\, celulele progenitoare continu\
s\ prolifereze, s\ se diferen]ieze [i s\ se matureze, conduc^nd la dou\ clase
mari de limfocite : limfocitele B [i limfocitele T. Capacitatea lor func]ional\
va asigura cooperarea, constituind elementele esen]iale ale sistemului
imunitar. Cele dou\ clase, de[i prea pu]in distincte morfologic, au func]ii
diferite [i, `n acela]i timp, exprim\ la nivel membranar o serie de structuri
antigenice care le va asigura func]ia [i recunoa[terea (imunofenotipaj).
Cele mai importante sunt dou\ tipuri de molecule cu func]ie de receptor :
imunglobulina mebranar\ exprimat\ de limfocitul B [i receptorul T (TCR T
cell receptor, cu molecula asociat\ CD3) exprimat de limfocitul T. Relativ
recent a fost identificat\ o a treia subpopula]ie limfocitar\, limfoctele non-B,
non-T sau limfocitele null.
Morfologie :
Limfocitele din s^nge sunt celule mici, `n raport cu celelalte elemente
figurate. Au un diametru de sub 10 microni. Excep]ie de la regul\ fac
limfocitele granulare mari care pot atinge un diametru dublu [i con]in `n
citoplasm\ granula]ii azurofile. Raportul nucleo-citoplasmatic este mare
cu tendin]\ c\tre 1. Nucleul este rotund-ovalar, de culoare `nchis\, cu o
cromatin\ grosier\, compactat\ `n gr\mezi. Nucleolii sunt invizibili.
Citoplasma formeaz\ o bordur\ discret\, abia vizibil\ (uneori) `n jurul
nucleului. Citoplasma este moderat bazofil\ [i, `n general, lipsit\ de
granula]ii.
Spre deosebire de eritrocite, granulocite [i trombocite, limfocitul
matur nu constituie elementul terminal al liniei sale, av^nd posibilitatea
ca sub o stimulare corespunz\toare s\-[i recapete capacit\]ile proliferative
cu reevolu]ie spre stadiul de limfoblast.
Limfoblastul are dimensiuni mai mari, cu diametrul `ntre 8 [i 12
microni, nucleul rotund/ovalar cu o cromatin\ mai fin\, lax\, cu nucleol
prezent, ocup^nd o treime din suprafa]a nuclear\. Citoplasma este mai
abundent\, bazofil\, bogat\ `n poliribozomi.
E. TROMBOCITOPOIEZA
Trombocitele
reprezint\
termenul
final
de
maturare
al
liniei
megakariocitare. Elementele acestei linii pot fi izolate din snge [i m\duva
osoas\ prin centrifugare sub gradient (ex Ficoll-hypaque), triaj celular cu
anticorpi monoclonali (Ac mo), [i cultivare pe medii semisolide n prezen]a
unui ser de la un pacient cu insuficien]\ medular\ cantitativ\ : serurile de la
astfel de pacien]i sunt bogate n factori de cre[tere.
Linia megakariocitar\ provine ca [i celelalte linii din celulele stem
hematopoietice (CSH). Linia ncepe n momentul n care celula
14
hematopoietic\ pluripotent\ se transform\ ntr-o celul\ capabil\ s\ prolifereze
[i s\ dea o colonie de megakariocite in vitro - progenitorul megacariocitar.
Dup\ un num\r de mitoze n compartimentul celulelor progenitoare, celula
ajunge ntr-un stadiu de tranzi]ie, n compartimentul celulelor precursoare,
n care celulele nu se mai divid, ele suferind doar endomitoze.
Compartimentul celulelor progenitoare este heterogen cu o ierarhie
precis\ constituind o linie continu\ de celule. Dou\ - trei tipuri de progenitori
au fost identificate fie pe baza studiului tipului de colonii ob]inut in vitro
(cu c^t m\rimea coloniei este mai mare sau cu c^t timpul s\u de apari]ie
`n cultur\ este mai lung, cu at^t progenitorul din care a provenit colonia
este mai primitiv), fie pe baza studiului ploidiei megakariocitelor ce
compun o colonie izolat\. Progenitorii megakariocitari se `mpart schematic
`n trei tipuri :
BFU-MK (Burst Forming Units - Megakaryocyte) - sunt progenitorii cei
mai primitivi. Dau colonii compuse din peste 50 celule organizate `n mai
multe subcolonii pe modelul coloniilor eritroblastice derivate din BFU-E.
Durata de apari]ie a coloniilor este de 12 zile la [oarece [i 21 zile la om.
Frecven]a lor `n m\duv\ este foarte slab\. Mai recent a fost descris un
progenitor denumit HPP (high proliferative potential)-CFU-MK care are
propriet\]i apropiate sau identice cu BFU-MK doar c\, colonia rezultat\
este grupat\ `ntr-o singur\ gr\mad\.
CFU-MK (Colony Forming Units Megacaryocyte) - aceste celule difer\ de
precedentele prin capacitarea proliferativ\ mai sc\zut\. Ele dezvolt\
colonii de megakariocite de 3 - 50 celule, `n 5 zile la [oarece [i 12 zile la
om. CFU-MK, spre deosebire de BFU-MK, sunt distruse dup\ un tratament
cu agen]i ca 5-fluoro-uracil, ceea ce determin\ ca recuperarea
trombocitar\ post-terapeutic\ s\ fie mai lent\ dec^t cea eritrocitar\.
LD-CFU-MK (Low density) - aceste celule au fost identificate la [oarece
ca un progenitor cu densitate celular\ sub 1.050 g/l. Ele dau, `n 2 3
zile, colonii formate dintr-un num\r mic de megakariocite cu ploidie
crescut\. Aceste celule corespund unui stadiu de diferen]iere de tranzi]ie
`n care, progenitorul megakariocitar `[i comut\ sistemul de proliferare
c\tre endomitoze.
Odat\ intrate n compartimentul precursorilor, sau de matura]ie,
celulele sufer\ un proces de matura]ie at^t nuclear ct [i citoplasmatic.
Din acest moment celula nu se mai divide. Nucleul [i dubleaz\ succesiv
con]inutul n ADN f\r\ a se divide (endomitoz\). Astfel celula [i cre[te
ploidia de la 2N pn\ la 64-128N. La om, majoritatea megakariocitelor sunt
16N (50%). Pe m\sur\ ce [i cre[te ploidia nucleul rotund devine anco[at
apoi polilobulat [i [i condenseaz\ cromatina. n paralel are loc o maturare a
citoplasmei cu formarea de granule: dense, alfa [i lizozomale, apari]ia unor
antigene plachetare, formarea memebranelor de demarca]ie ale
sistemului de membrane ce comunic\ cu exteriorul. Ele provin din
invaginarea membranei plasmatice. Au rolul de a constitui o rezerv\ de
membrane a megakariocitelor. Astfel, `nainte de a elibera trombocitele,
megakariocitul se alunge[te lu^nd forma unei caracati]e, alungire ce se
face datorit\ rezervei de membrane [i derul\rii ei cu ajutorul microtubilor.
15
La intervale regulate apar constric]ii unde se concentreaz\ microtubii [i
microfilamentele care vor rupe citoplasma [i vor forma trombocitele.
n aceast compartiment megakariocitul trece prin mai multe stadii :
Stadiul I (megakarioblast) - prima celul\ recognoscibil\ morfologic a
seriei. Este o celul\ blastic\ cu diametrul de 15-50 microni. Raportul
nucleo-citoplasmatic este mare. Nucleul este mare, ovalar sau anco[at
(form\ de rinichi) cu nucleoli eviden]i (2-6). Citoplasma este bazofil\, f\r\
granula]ii ;
Stadiul II (promegakariocit, megakariocitul bazofil) - este o celul\ mai
mare 20-80 microni cu nucleu polilobat [i citoplasma policromatofil\,
mai abundent\ [i con]innd granula]ii azurofile a[ezate perinuclear.
Stadiul III (megakariocitul granular) - celul\ foarte mare, cu diametrul de
35-160 microni, cu un nucleu neregulat, polilobat compus din cromatin\
dens\, f\r\ nucleoli vizibili. Citoplasma este abundent\, albastru deschis
cu granula]ii azurofile la periferie.
Megakariopoieza are loc la nivelul m\duvii osoase unde
megakariocitele se reg\sesc preferen]ial la nivelul subendoteliului
sinusoidelor. Fiecare megakariocit d\ na[tere la 1.000-8.000 de plachete.
Se discut\ tot mai mult rolul pl\mnului ca situs principal de producere a
plachetelor (din megakariocite ce au p\r\sit m\duva prin pasaj
transendotelial, [i au fost captate la acest nivel). n ceea ce prive[te
mecanismul de producere a plachetelor exist\ 4 teorii :
plachetele ar nmuguri la suprafa]a megakariocitelor - foarte
pu]in probabil
teritoriile plachetare ar fi predeterminate ca m\rime [i form\ n
interiorul megakariocitelor ;
megakariocitele s-ar fragmenta n pl\mn sub efectul for]elor
fizice ale curentului sanguin ;
teoria
cea
mai acceptat\
este
aceea
potrivit
c\reia
megakariocitele ar trimite prelungiri lungi (pseudopode) care se
fragmenteaz\ n plachete. Fragmentarea ar avea loc n circuitul
sanghin unde a fost trimis pseudopodul (prin endoteliul vascular).
c. Reglarea tombocitopoiezei
Reglarea produc]iei de plachete se realizeaz\ prin intermediul a trei
parametri independen]i :
num\rul de megakariocite (mecanismul de proliferare), volumul
mgakariocitelor;
talia megakariocitelor care depinde de ploidia lor. Cre[terea ploidiei
rezult\ prin endomitoz\, `nso]it\ de cre[terea volumului celular [i
citoplamatic.
maturarea citoplasmatic\ (cu dezvoltarea re]elei membranare
intracitoplasmatice membrane de demarca]ie care vor servi ca
membrate citoplasmatice pentru trombocite).
Reglarea produc]iei plachetare (megakariopoieza), conform ipotezi
dublei regl\ri survine la dou\ niveluri celulare distincte : o prim\ citokin\
(MK-CSF) ac]ioneaz\ numai asupra etapelor precoce ale diferen]ierii
(celulele progenitoare), [i un factor tardiv de diferen]iere (trombopoietina)
ce influen]eaz\ procesul de maturare al megakariocitelor [i ploidizarea lor.
16
Cercet\rile au ar\tat c\ IL-3 [i GM-CSF joac\ rolul de MK-CSF fiind capabile s\
induc\ formarea de colonii megakariocitare prin ac]iunea concomitent\
asupra BFU-MK [i CFU-MK. Trombopoietina ac]ioneaz\ asupra etapelor
ulterioare al\turi de citokinele din familia IL-6. Pe de alt\ parte,
megakariopoieza este inhibat\ n mod fiziologic de factori inhibitori
elibera]i de plachetele activate: TGF-beta, PF4, beta tromboglobulina.
Trombopoietina (TPO) este hormonul fiziologic care regleaz\
produc]ia trombocitar\. Studiile au demonstrat faptul c\, pe l^ng\ ac]iunea
sa preponderant\ asupra liniei megakariocitare, TPO ar juca un rol principal
`n etapele precoce ale hematopoiezei.
Gena TPO uman\ este situat\ pe cromozomul 3, `n 3q27-28.. Gena
este exprimat\ `n mod esen]ial, `n ficat, rinichi dar [i `n m\duva osoas\, de
fibroblastele medulare `n cultur\ [i de celulele endoteliale din cordonul
ombilical.
In vitro, TPO este responsabil\ de :
cre[terea taliei [i ploidiei (deobicei peste 64 N) a megakariocitelor
(MK) `n cultur\;
inducerea form\rii de colonii megacariocitare de talie mic\,
comport^nd 3 - 5 MK pe colonie;
inducerea de markeri de diferen]iere megakariocitar\ pe celulele
CD34+ : glicoproteina GP Ib et glicoproteina GP IIb/IIIa.
17
S-ar putea să vă placă și
- Curs 1.hematopoiezaDocument47 paginiCurs 1.hematopoiezanina.aÎncă nu există evaluări
- Curs 1 HematopiezaDocument8 paginiCurs 1 Hematopiezastefanpaun100% (1)
- Hematopoieza NormalaDocument7 paginiHematopoieza NormalaSmeu-Mare Mihai-Alexandru100% (1)
- Sange Hematopoeza AmalineiDocument20 paginiSange Hematopoeza AmalineiEduard CarpÎncă nu există evaluări
- 9.sindroamele HemoragipareDocument9 pagini9.sindroamele HemoragipareMadalina SercaianuÎncă nu există evaluări
- Curs 1 Facultate 16.10.2014Document33 paginiCurs 1 Facultate 16.10.2014Ioana-Mihaela TomaÎncă nu există evaluări
- Suport Curs HematopoiezaDocument14 paginiSuport Curs HematopoiezaANDREEA FLOREAÎncă nu există evaluări
- Hematologie Clinica Curs Dat La Farmacie An IIDocument129 paginiHematologie Clinica Curs Dat La Farmacie An IIManuela Stefu100% (1)
- Hematopoieza 2010Document61 paginiHematopoieza 2010Oana AldeaÎncă nu există evaluări
- I. HematopoiezaDocument17 paginiI. HematopoiezaAna Maria ȘuteuÎncă nu există evaluări
- Determinarea HematocrituluiDocument4 paginiDeterminarea HematocrituluiCojocariu Claudiu100% (2)
- 01 SângeleDocument38 pagini01 SângeleCatalin StanciulescuÎncă nu există evaluări
- LP 1 Fiziologie FARMACIE An 1 Sem 2Document62 paginiLP 1 Fiziologie FARMACIE An 1 Sem 2Mihaila LenutaÎncă nu există evaluări
- Numarare LeucociteDocument4 paginiNumarare LeucociteMirea Silvian100% (1)
- Histologie SangeleDocument11 paginiHistologie Sangeleiulia-uroÎncă nu există evaluări
- Parazitologie Complet PDFDocument12 paginiParazitologie Complet PDFNeli Mihaela100% (1)
- Hemogram ADocument36 paginiHemogram AUngureanu AndreiÎncă nu există evaluări
- Cuprins Compendiu de HematologieDocument1 paginăCuprins Compendiu de Hematologiediddi21100% (1)
- Cinetica EritrocitelorDocument42 paginiCinetica EritrocitelorOana Rosca100% (1)
- Tematica Medicina de Laborator 2022Document5 paginiTematica Medicina de Laborator 2022madalina apostolÎncă nu există evaluări
- Anemiile IDocument66 paginiAnemiile IMihaela FloreaÎncă nu există evaluări
- 8.sangele Periferic - Suport Curs - 2021Document17 pagini8.sangele Periferic - Suport Curs - 2021CIPSITÎncă nu există evaluări
- HematoDocument47 paginiHematoCarmen Maria100% (2)
- BABYDocument651 paginiBABYmoni_ignatÎncă nu există evaluări
- Curs NR 1 HematologieDocument12 paginiCurs NR 1 HematologieAna HancÎncă nu există evaluări
- Curs 4 - Trombocite, HemostazaDocument14 paginiCurs 4 - Trombocite, HemostazaCiprian Rata100% (1)
- Hemograma-Leucemia Granulocitara CronicaDocument29 paginiHemograma-Leucemia Granulocitara CronicaAnonymous pPacR7ydB100% (1)
- Determinarea Timpilor de CoagulareDocument12 paginiDeterminarea Timpilor de CoagulareAdrian Pavelescu100% (1)
- 1.anemiile HemoliticeDocument7 pagini1.anemiile HemoliticeMadalina Sercaianu100% (1)
- Subiecte Biochimie LP 2Document21 paginiSubiecte Biochimie LP 2CristiÎncă nu există evaluări
- Explorarea HemostazeiDocument51 paginiExplorarea Hemostazeianidale100% (1)
- 1 - Curs HEMATOPOIEZA - DG ANEMIIDocument101 pagini1 - Curs HEMATOPOIEZA - DG ANEMIIAlinaÎncă nu există evaluări
- 63343478-Hemograma pdf1Document26 pagini63343478-Hemograma pdf1Alexuta MariaÎncă nu există evaluări
- CoagulareDocument127 paginiCoagulareRaf AlkÎncă nu există evaluări
- MVP - Laborator 2Document7 paginiMVP - Laborator 2Lucian BaguÎncă nu există evaluări
- Cursuri-Barem Hematologie BadeaDocument80 paginiCursuri-Barem Hematologie BadeaMaria Dinu100% (1)
- Curs 4Document63 paginiCurs 4Ioana CÎncă nu există evaluări
- 13.curs Oncogeneza Markeri Tumorali25.05Document65 pagini13.curs Oncogeneza Markeri Tumorali25.05popoviciÎncă nu există evaluări
- Aspecte Normale Ale EritrocitelorDocument1 paginăAspecte Normale Ale EritrocitelorAndriucFlaviaÎncă nu există evaluări
- HematopoiezaDocument51 paginiHematopoiezaNico Ionaşcu100% (1)
- Trombocite - Ce Inseamna Trombocite Scazute Sau Crescute - Cancer360 PDFDocument5 paginiTrombocite - Ce Inseamna Trombocite Scazute Sau Crescute - Cancer360 PDFMarcelApostolacheÎncă nu există evaluări
- IMUNOFENOTIPAREDocument29 paginiIMUNOFENOTIPAREIoana VoinescuÎncă nu există evaluări
- Curs Gamapatii Monoclonale2Document31 paginiCurs Gamapatii Monoclonale2Daniela GrigÎncă nu există evaluări
- Eritrocitele+Formula LeucocitaraDocument14 paginiEritrocitele+Formula LeucocitaraMonica Ispas100% (1)
- HematiiDocument53 paginiHematiiOana DrăganÎncă nu există evaluări
- LP Sange 1 - Fiziologia SangeluiDocument21 paginiLP Sange 1 - Fiziologia SangeluiAnghel Ana-MariaÎncă nu există evaluări
- Biochimia Hemostazei (From Biochimie Clinica)Document25 paginiBiochimia Hemostazei (From Biochimie Clinica)MedlibraryÎncă nu există evaluări
- CURS Sindroame HemoragiceDocument60 paginiCURS Sindroame HemoragiceRaluca MirceaÎncă nu există evaluări
- Diagnostic de OrganDocument4 paginiDiagnostic de OrganDitescu Beatrice-AleqsandraÎncă nu există evaluări
- CONTEC EKG - Monitoare 02 (1) .2019 1 db-1Document8 paginiCONTEC EKG - Monitoare 02 (1) .2019 1 db-1Raluca CotosÎncă nu există evaluări
- ANEMIIDocument22 paginiANEMIIStefania NicolaeÎncă nu există evaluări
- TromboliticeDocument25 paginiTromboliticezsrenaÎncă nu există evaluări
- Hematologie MG V Hematopoieza Si Anemii PT Test Martie 2013Document34 paginiHematologie MG V Hematopoieza Si Anemii PT Test Martie 2013Carmen JurcoaneÎncă nu există evaluări
- Curs 5 Hematopoieza Grupe SangeDocument81 paginiCurs 5 Hematopoieza Grupe SangeMischa Vlăsceanu100% (1)
- 6 HematopoezaDocument8 pagini6 HematopoezaValentin MoisăÎncă nu există evaluări
- Introducere in Hematologia Clinica BTDocument358 paginiIntroducere in Hematologia Clinica BTDidiÎncă nu există evaluări
- Hematopoieza - RDocument36 paginiHematopoieza - RStarks GazerÎncă nu există evaluări
- Hematologie Clinica ROMANA Pus Pe IntranetDocument168 paginiHematologie Clinica ROMANA Pus Pe IntranetIoana PietricicaÎncă nu există evaluări
- 2.b. Curs 2 Imunologie 2021 (Hematopoieza)Document67 pagini2.b. Curs 2 Imunologie 2021 (Hematopoieza)Cristian StanÎncă nu există evaluări
- HematopoezaDocument4 paginiHematopoezaBogdan DianaÎncă nu există evaluări
- Neuropatia DiabeticaDocument31 paginiNeuropatia DiabeticaTeona OanaÎncă nu există evaluări
- Prezentare CerebelosDocument28 paginiPrezentare CerebelosTeona OanaÎncă nu există evaluări
- Vi. Nervul AbducensDocument3 paginiVi. Nervul AbducensTeona OanaÎncă nu există evaluări
- Codul de Etica Si Deontologie Al Asistentului MedicalDocument10 paginiCodul de Etica Si Deontologie Al Asistentului MedicalTeona OanaÎncă nu există evaluări
- Prezentare de Caz. EpilepsieDocument37 paginiPrezentare de Caz. EpilepsieTeona OanaÎncă nu există evaluări
- Sindromul PiramidalDocument26 paginiSindromul PiramidalTeona OanaÎncă nu există evaluări
- 15 Boala ParkinsonDocument48 pagini15 Boala ParkinsonElena ElenaaÎncă nu există evaluări
- Tremor ms1Document34 paginiTremor ms1Teona OanaÎncă nu există evaluări
- Neuro LogieDocument1 paginăNeuro LogieTeona OanaÎncă nu există evaluări