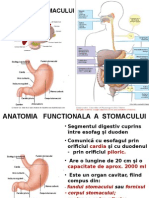
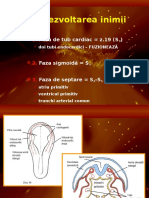
S-ar putea să vă placă și
- Antoine de Saint-Exupéry - Micul Prinţ PDFDocument43 paginiAntoine de Saint-Exupéry - Micul Prinţ PDFCris Postolache100% (1)
- Revolutia CardiacaDocument78 paginiRevolutia CardiacaCristinaÎncă nu există evaluări
- Tesutul EpitelialDocument6 paginiTesutul EpitelialAriela Săvuţ100% (7)
- Subiecte Anatomie An II Sem I, Partea IDocument39 paginiSubiecte Anatomie An II Sem I, Partea IAndrei Manea100% (2)
- Suport Curs Perform Mecanograme 2018Document127 paginiSuport Curs Perform Mecanograme 2018Ronaldo LazarÎncă nu există evaluări
- Subiecte Colocviu LP MD 2016-2017 Sem IDocument1 paginăSubiecte Colocviu LP MD 2016-2017 Sem IMdÎncă nu există evaluări
- Ciclul Cardiac - Mecanograme.Document9 paginiCiclul Cardiac - Mecanograme.Stefan PaunÎncă nu există evaluări
- Genetica CursDocument4 paginiGenetica CursRonaldo LazarÎncă nu există evaluări
- Genetica CursDocument4 paginiGenetica CursRonaldo LazarÎncă nu există evaluări
- TalamusDocument6 paginiTalamusGabrielleÎncă nu există evaluări
- Sinteza - Sistemul MuscularDocument11 paginiSinteza - Sistemul Muscularlesnoland100% (1)
- Regiunea NucalaDocument4 paginiRegiunea NucalaFlavius Ion NicolaeÎncă nu există evaluări
- Bio Cel Curs 6 Transportul TransmembranarDocument54 paginiBio Cel Curs 6 Transportul TransmembranarMarin TheodorÎncă nu există evaluări
- Test General de BiologieDocument5 paginiTest General de BiologieAvillaÎncă nu există evaluări
- 1echilibrul OsmoticDocument44 pagini1echilibrul OsmoticRachel MNÎncă nu există evaluări
- Transmiterea Sinaptica L Zagrean UmfcdDocument17 paginiTransmiterea Sinaptica L Zagrean UmfcdRoxana Androne50% (2)
- LP Ciclul Cardiac - ZgomoteDocument10 paginiLP Ciclul Cardiac - ZgomoteAndrei RaduÎncă nu există evaluări
- Farmacologie - Farmacologia Sistemului Nervos ParasimpaticDocument14 paginiFarmacologie - Farmacologia Sistemului Nervos ParasimpaticMoNistorÎncă nu există evaluări
- Subiecte Colocviu Biochimie 2020Document3 paginiSubiecte Colocviu Biochimie 2020Ariana-Teodora PotoracÎncă nu există evaluări
- MB Inf VascularizaţieDocument67 paginiMB Inf VascularizaţieDragos AlexandruÎncă nu există evaluări
- Curs 4 SemIIDocument12 paginiCurs 4 SemIIoanaobrejaÎncă nu există evaluări
- Embriologie LP 1, An I, Sem. IIDocument43 paginiEmbriologie LP 1, An I, Sem. IITed Kami-samaÎncă nu există evaluări
- Nucleul - Notite de CursDocument19 paginiNucleul - Notite de CursMorgan Rosewood100% (1)
- Trauma Atlas RezidențiatDocument26 paginiTrauma Atlas RezidențiatRonaldo LazarÎncă nu există evaluări
- Farmacologie Curs 7Document13 paginiFarmacologie Curs 7MarcNicaÎncă nu există evaluări
- CarotidogramaDocument3 paginiCarotidogramaAromadulce100% (1)
- Degradarea Scheletului Atomilor de CarbonDocument11 paginiDegradarea Scheletului Atomilor de CarbonCatalin Apetrei100% (2)
- C5 Distensibilitatea Vasculara Si Functiile Sistemelor Arterial Si VenosDocument4 paginiC5 Distensibilitatea Vasculara Si Functiile Sistemelor Arterial Si VenosEphebus EfebÎncă nu există evaluări
- Curs 1 Boala de Reflux Gastro-EsofagianDocument22 paginiCurs 1 Boala de Reflux Gastro-Esofagianflorentintoc707Încă nu există evaluări
- Proprietati AdnDocument8 paginiProprietati AdnBada BanuÎncă nu există evaluări
- Fiziologia Muschiului - 2019Document43 paginiFiziologia Muschiului - 2019Narcis MessiÎncă nu există evaluări
- Benevron B ProspectDocument5 paginiBenevron B ProspectvasiliklÎncă nu există evaluări
- Curs 9 Metabolismul BilirubineiDocument32 paginiCurs 9 Metabolismul BilirubineiTotolici StefanÎncă nu există evaluări
- Carbonic AnhidrazaDocument7 paginiCarbonic AnhidrazacroitorugeorgianaÎncă nu există evaluări
- Lp4-Specializarile Plasmalemei PDFDocument5 paginiLp4-Specializarile Plasmalemei PDFOvidiu MarianÎncă nu există evaluări
- T1Document12 paginiT1Vasilică SavaÎncă nu există evaluări
- Fiziologie Si Patologie-ColocviuDocument25 paginiFiziologie Si Patologie-ColocviugencimoravaÎncă nu există evaluări
- Formule BiofizicaDocument4 paginiFormule BiofizicaMihai HondruÎncă nu există evaluări
- Sistemul ComplementDocument25 paginiSistemul ComplementanairamÎncă nu există evaluări
- Hemofilia Sau Boala RegilorDocument3 paginiHemofilia Sau Boala RegilorCristinaZgardanÎncă nu există evaluări
- Anatomie Sem. 2 - Curs 2Document4 paginiAnatomie Sem. 2 - Curs 2Andreea SchmitzerÎncă nu există evaluări
- Fiziologie Curs - HemodinamicaDocument7 paginiFiziologie Curs - HemodinamicaCiprian DolhescuÎncă nu există evaluări
- Anatomia Morfofuncțională Și Clinică A RinichilorDocument18 paginiAnatomia Morfofuncțională Și Clinică A RinichilorManta VladÎncă nu există evaluări
- Examenul Fizic PulmonarDocument32 paginiExamenul Fizic PulmonarcnajesrnasiferfÎncă nu există evaluări
- Glandele SuprarenaleDocument6 paginiGlandele SuprarenaleCristiana VrancuțaÎncă nu există evaluări
- SEMIOLOGIE Curs AP - RenalDocument4 paginiSEMIOLOGIE Curs AP - RenalgabiportuÎncă nu există evaluări
- Curs 1-Imunogenetica Si Transplantul - ComplexulDocument33 paginiCurs 1-Imunogenetica Si Transplantul - ComplexulCristian EneÎncă nu există evaluări
- Curs 1 Cergan An 2Document92 paginiCurs 1 Cergan An 2GEORGIANA-CIPRIANA CIUBOTARIUÎncă nu există evaluări
- Modificări Post-TranscriereDocument36 paginiModificări Post-TranscriereAna-Maria IonescuÎncă nu există evaluări
- Nervul Trigemen AnatomieDocument14 paginiNervul Trigemen AnatomieDunas DanÎncă nu există evaluări
- C8 Aparatul DigestivDocument46 paginiC8 Aparatul DigestivTheodora IrinaÎncă nu există evaluări
- LP 2 Analiza Cromozomilor Sexuali La OmDocument3 paginiLP 2 Analiza Cromozomilor Sexuali La OmAna Maria DanÎncă nu există evaluări
- Excretia 2Document10 paginiExcretia 2Alex_Burn_99130% (1)
- Referat docxVIOLETADocument10 paginiReferat docxVIOLETAtuturici liviuÎncă nu există evaluări
- LP 6Document4 paginiLP 6Laura Florea100% (1)
- Teratomul OvarianDocument12 paginiTeratomul OvarianPatrascu CristiÎncă nu există evaluări
- CCM2Document19 paginiCCM2Melnic CristinaÎncă nu există evaluări
- MicrocirculatiaDocument6 paginiMicrocirculatiaCiprian DolhescuÎncă nu există evaluări
- Tromboembolia Acută A Vaselor MezenterialeDocument76 paginiTromboembolia Acută A Vaselor MezenterialeElla LenusikÎncă nu există evaluări
- Grile 2014 CraiovaDocument62 paginiGrile 2014 CraiovaElinaÎncă nu există evaluări
- Genetica Si CancerulDocument12 paginiGenetica Si CancerulElena CaranicaÎncă nu există evaluări
- Insuficienta Renala AcutaDocument8 paginiInsuficienta Renala AcutaBeatrice StefanescuÎncă nu există evaluări
- 5 Anomalii Ale DiurezeiDocument18 pagini5 Anomalii Ale DiurezeiCocosul Cocosului CocosaruluiÎncă nu există evaluări
- Subiecte Examen CardiologieDocument7 paginiSubiecte Examen Cardiologiemikidog1Încă nu există evaluări
- Curs 7 Tesutul Osos PPT PDFDocument45 paginiCurs 7 Tesutul Osos PPT PDFAlex AlxÎncă nu există evaluări
- Anatomia Si Fiziologia Aparatului Sexual Masculin Si FemininDocument17 paginiAnatomia Si Fiziologia Aparatului Sexual Masculin Si FemininCristina AmbrusÎncă nu există evaluări
- Curs 12 Anatomia Si Fiziologia Aparatului Sexual Masculin Si FemininDocument21 paginiCurs 12 Anatomia Si Fiziologia Aparatului Sexual Masculin Si FemininMinerva TutoveanuÎncă nu există evaluări
- Sistem Genital Masculin Amalinei PDFDocument11 paginiSistem Genital Masculin Amalinei PDFMihai HondruÎncă nu există evaluări
- 22.sistemul Reproducator MasculinDocument16 pagini22.sistemul Reproducator MasculinioanaÎncă nu există evaluări
- Organele de Reproducere BarbatestiDocument7 paginiOrganele de Reproducere BarbatestiGeanina PavelÎncă nu există evaluări
- 03 HipoglicemiileDocument3 pagini03 HipoglicemiileSorin IoacaraÎncă nu există evaluări
- Regulament Senat 13.10.15Document27 paginiRegulament Senat 13.10.15Ronaldo LazarÎncă nu există evaluări
- TEMATICA - REZIDENTIAT - 2021 - G6 (Modificat)Document3 paginiTEMATICA - REZIDENTIAT - 2021 - G6 (Modificat)Andreea DașoveanuÎncă nu există evaluări
- Curs PZ 2 FlagelatDocument11 paginiCurs PZ 2 FlagelatRonaldo LazarÎncă nu există evaluări
- Regulament Alegere Studenti Reprezentanti Facultatea de MedicinaDocument12 paginiRegulament Alegere Studenti Reprezentanti Facultatea de MedicinaDragos RoscaÎncă nu există evaluări
- Metodologie Privind Acordarea BurselorDocument40 paginiMetodologie Privind Acordarea BurselorMaria DamocÎncă nu există evaluări
- Redactare AcademicaDocument1 paginăRedactare AcademicaRonaldo Lazar0% (1)
- CD Leziuni Elementare - OsDocument42 paginiCD Leziuni Elementare - OsStefanÎncă nu există evaluări
- Coco BaciliDocument5 paginiCoco BaciliRonaldo LazarÎncă nu există evaluări
- Buraga RenalDocument53 paginiBuraga RenalRonaldo LazarÎncă nu există evaluări
- Regulament Alegere Studenti Reprezentanti Facultatea de MedicinaDocument12 paginiRegulament Alegere Studenti Reprezentanti Facultatea de MedicinaDragos RoscaÎncă nu există evaluări
- Curs PZ 2 FlagelatDocument11 paginiCurs PZ 2 FlagelatRonaldo LazarÎncă nu există evaluări
- Subiectul 2 Anatomie An 2 Sem 2Document1 paginăSubiectul 2 Anatomie An 2 Sem 2Ronaldo LazarÎncă nu există evaluări
- Curs Histologie Polaritatea Membranei CelulareDocument12 paginiCurs Histologie Polaritatea Membranei CelulareRonaldo LazarÎncă nu există evaluări
- Cavitatea BucalaDocument5 paginiCavitatea BucalaametisÎncă nu există evaluări
- Embriologie Sistem CardiovascularDocument27 paginiEmbriologie Sistem CardiovascularStrumphiÎncă nu există evaluări
- Primul Subiect La Anato Sem 2 An 2Document1 paginăPrimul Subiect La Anato Sem 2 An 2Ronaldo LazarÎncă nu există evaluări
- Rezumatul Tezei de Doctorat Cella Serghi-Studiu MonograficDocument10 paginiRezumatul Tezei de Doctorat Cella Serghi-Studiu MonograficRonaldo LazarÎncă nu există evaluări
- Centrul Judeţean de Conservare Şi Valorificare A Tradiţiei Şi Creaţiei PopulareDocument1 paginăCentrul Judeţean de Conservare Şi Valorificare A Tradiţiei Şi Creaţiei PopulareRonaldo LazarÎncă nu există evaluări