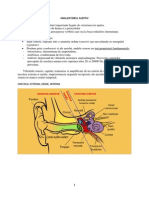
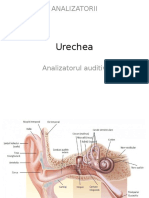
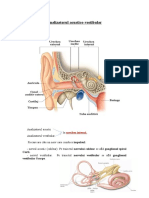
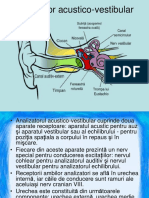
S-ar putea să vă placă și
- BIOACUSTICADocument8 paginiBIOACUSTICAKasai Maria Petronela100% (3)
- Titlul Lucrarii - Organul de Simt - UrecheaDocument5 paginiTitlul Lucrarii - Organul de Simt - UrecheaDaniel NicolaeÎncă nu există evaluări
- Analizatorul Acustic Si Analizatorul VestibularDocument8 paginiAnalizatorul Acustic Si Analizatorul VestibularDragosh Ionut ProtopopescuÎncă nu există evaluări
- Sunetul fizicii: Acustica fenomenologicăDe la EverandSunetul fizicii: Acustica fenomenologicăEvaluare: 4.5 din 5 stele4.5/5 (2)
- Biofizica Receptiei AuditiveDocument14 paginiBiofizica Receptiei AuditiveAndrone Camelia100% (1)
- Anatomia Si Fiziologia Analizatorului AuditivDocument7 paginiAnatomia Si Fiziologia Analizatorului AuditivAnabela Alexandra100% (1)
- Analizatorul AuditivDocument11 paginiAnalizatorul AuditivAlexandra100% (2)
- Analizatorul Acustico VestibularDocument5 paginiAnalizatorul Acustico Vestibularandreea teodoraÎncă nu există evaluări
- Biofizica Analizatorilor FiziciDocument45 paginiBiofizica Analizatorilor FiziciRadu NoțingherÎncă nu există evaluări
- URECHEADocument25 paginiURECHEAToma GeorgianaaÎncă nu există evaluări
- Biofizica Receptiei AuditiveDocument18 paginiBiofizica Receptiei AuditiveIoana Mihaela SerbanÎncă nu există evaluări
- Biofizica Recepţiei AuditiveDocument19 paginiBiofizica Recepţiei AuditiveMihai ArcaleanÎncă nu există evaluări
- Biofizica Sistemelor ComplexeDocument9 paginiBiofizica Sistemelor ComplexeIulia PopÎncă nu există evaluări
- Aparatul Auditiv Si VizualDocument8 paginiAparatul Auditiv Si VizualAndra Apostol100% (1)
- Analizatorul AuditivDocument7 paginiAnalizatorul AuditivSmadu AdrianaÎncă nu există evaluări
- 8.fiziologia Aparatului de Transmitere A SunetelorDocument3 pagini8.fiziologia Aparatului de Transmitere A SunetelorAndrei RotaruÎncă nu există evaluări
- Analizatorul AuditivDocument15 paginiAnalizatorul Auditivcrr_29Încă nu există evaluări
- Receptia AuditivaDocument31 paginiReceptia AuditivaSebastian StoicaÎncă nu există evaluări
- UrecheaDocument6 paginiUrecheaTeodor Mihai100% (2)
- Biomecanica Sistemului AuditivDocument8 paginiBiomecanica Sistemului AuditivStefania FrincuÎncă nu există evaluări
- BIOACUSTICADocument4 paginiBIOACUSTICAmonicaudreaÎncă nu există evaluări
- Biofizica Recepţiei AuditiveDocument22 paginiBiofizica Recepţiei AuditiveValentino PetreÎncă nu există evaluări
- Analizatorul Acustic Si VestibularDocument31 paginiAnalizatorul Acustic Si VestibularLaura GareaÎncă nu există evaluări
- Receptia AuditivaDocument10 paginiReceptia AuditivaRadu BulboacaÎncă nu există evaluări
- BIOACUSTICADocument8 paginiBIOACUSTICAkonadelle4390100% (1)
- SunetulDocument15 paginiSunetulcorinaburlacu110Încă nu există evaluări
- Biofizica Receptiei AuditiveDocument6 paginiBiofizica Receptiei AuditiveIulia Vasilica GirosanÎncă nu există evaluări
- Proiect Urechea BionicaDocument7 paginiProiect Urechea BionicaFlorin PîrîialăÎncă nu există evaluări
- 4 Urechea-REFERATDocument5 pagini4 Urechea-REFERATDavid AndreiÎncă nu există evaluări
- Aparatul AuditivDocument10 paginiAparatul AuditivCorina FintaÎncă nu există evaluări
- Analizatorul AcusticoDocument15 paginiAnalizatorul AcusticoOana Stefania PintiliiÎncă nu există evaluări
- Analizatorul AuditivDocument18 paginiAnalizatorul AuditiveugenalexandrovÎncă nu există evaluări
- Analizatorul Auditiv Rol Si Cractere GeneraleDocument11 paginiAnalizatorul Auditiv Rol Si Cractere GeneraleIRINA-MIHAELA TURCUÎncă nu există evaluări
- Analizatorul AuditivDocument10 paginiAnalizatorul AuditivMiron EvelinaÎncă nu există evaluări
- Analizatorul AuditivDocument5 paginiAnalizatorul AuditivCristina LoveÎncă nu există evaluări
- Analizator AuditivDocument23 paginiAnalizator AuditivHaz Adiana100% (2)
- Biofizica Analizatorului Acustic: Notiuni de Anatomie Si FiziologieDocument6 paginiBiofizica Analizatorului Acustic: Notiuni de Anatomie Si FiziologieHoratiu Ioan MateiuÎncă nu există evaluări
- Profesionisti Deficienta Auditiva RuDocument92 paginiProfesionisti Deficienta Auditiva RuLaurentia AvramÎncă nu există evaluări
- Analizatorul Auditiv La OmDocument11 paginiAnalizatorul Auditiv La OmIon CroitoruÎncă nu există evaluări
- Cum AuzimDocument19 paginiCum AuzimAlexandru ClisuÎncă nu există evaluări
- 3.3 Caile AuditiveDocument2 pagini3.3 Caile AuditiveMarinela MureaÎncă nu există evaluări
- Auditivsubiectul 3Document3 paginiAuditivsubiectul 3Russu DumitruÎncă nu există evaluări
- Fiziologia Sistemului AuditivDocument20 paginiFiziologia Sistemului AuditivOana Cristina100% (1)
- Organul AuzuluiDocument21 paginiOrganul AuzuluiLilia LapteacruÎncă nu există evaluări
- Curs 4 Bioacustica AP - AuditivDocument7 paginiCurs 4 Bioacustica AP - AuditivIulia Vasilica GirosanÎncă nu există evaluări
- Vestib AuditivDocument22 paginiVestib AuditivLiubovi CuharencoÎncă nu există evaluări
- Lumea SunetelorDocument11 paginiLumea SunetelorCristina Maria DeneşanÎncă nu există evaluări
- LihjDocument74 paginiLihjanushka1988Încă nu există evaluări
- Anatomia, Fiziologia, Patologia Organelor de Auz, Văz Și Vorbire PDFDocument74 paginiAnatomia, Fiziologia, Patologia Organelor de Auz, Văz Și Vorbire PDFanushka1988Încă nu există evaluări
- UrecheaDocument22 paginiUrechearaysa_svÎncă nu există evaluări
- XI - Biologia (A. 2020, in Limba Romana) - 32-35Document4 paginiXI - Biologia (A. 2020, in Limba Romana) - 32-35Diana GuzunÎncă nu există evaluări
- Analizatorul Acustico VestibularDocument7 paginiAnalizatorul Acustico VestibularZoeÎncă nu există evaluări
- Texte de Prelegeri - Anatomia Și Fiziologia Sistemelor Senzoriale (Organelor de Auz, Văz, Vorbire)Document74 paginiTexte de Prelegeri - Anatomia Și Fiziologia Sistemelor Senzoriale (Organelor de Auz, Văz, Vorbire)natalia silivestruÎncă nu există evaluări
- Notiuni de Anatomie Si Fizionomie A UrechiiDocument15 paginiNotiuni de Anatomie Si Fizionomie A UrechiiCrina Maria PuiaÎncă nu există evaluări
- Serban Andreea MariaDocument6 paginiSerban Andreea MariaHucÎncă nu există evaluări
- Mecanoreceptia La VertebrateDocument8 paginiMecanoreceptia La VertebrateCamelia ManeaÎncă nu există evaluări
- LP 12 Sângele Și HematopoiezaDocument18 paginiLP 12 Sângele Și HematopoiezaSergiuR94Încă nu există evaluări
- LP 7 Țesuturi ConducătoareDocument18 paginiLP 7 Țesuturi ConducătoareSergiuR94Încă nu există evaluări
- Lp. 4 MicroDocument4 paginiLp. 4 MicroSergiuR94Încă nu există evaluări
- Lp. 1 MicroDocument9 paginiLp. 1 MicroSergiuR94Încă nu există evaluări
- LP 7 Peretele Celular Si Permeabilitatea CelularăDocument43 paginiLP 7 Peretele Celular Si Permeabilitatea CelularăSergiuR94100% (1)
- Curs 8 9 AnorganicaDocument18 paginiCurs 8 9 AnorganicaSergiuR94Încă nu există evaluări
- Biologie Celulară - LP - 5Document37 paginiBiologie Celulară - LP - 5SergiuR94Încă nu există evaluări
- C 9 - Organizarea-Si-Exprimarea-Materialului-Genetic-La-ProcarioteDocument24 paginiC 9 - Organizarea-Si-Exprimarea-Materialului-Genetic-La-ProcarioteSergiuR94100% (1)
- Biologie Celulară - LP - 3Document19 paginiBiologie Celulară - LP - 3SergiuR94Încă nu există evaluări
- LP 8 Celula AnimalăDocument18 paginiLP 8 Celula AnimalăSergiuR94Încă nu există evaluări
- LP 5 NucleulDocument35 paginiLP 5 NucleulSergiuR94Încă nu există evaluări
- LP 3 Principalii Constituenți CelulariDocument10 paginiLP 3 Principalii Constituenți CelulariSergiuR94Încă nu există evaluări
- LP 4 Organite CitoplasmaticeDocument22 paginiLP 4 Organite CitoplasmaticeSergiuR94Încă nu există evaluări
- C 12 - Replicarea Şi Biosinteza ADN. Transcrierea Mesajului Genetic. Traducerea Codului Genetic Şi Biosinteza Proteinelor La Eucariote. Mecanisme Reparatorii ADNDocument23 paginiC 12 - Replicarea Şi Biosinteza ADN. Transcrierea Mesajului Genetic. Traducerea Codului Genetic Şi Biosinteza Proteinelor La Eucariote. Mecanisme Reparatorii ADNSergiuR94Încă nu există evaluări
- C 13 - Moartea Celulară Programată Genetic Apoptoza. Senescenţa Celulară, Necroză.Document15 paginiC 13 - Moartea Celulară Programată Genetic Apoptoza. Senescenţa Celulară, Necroză.SergiuR94Încă nu există evaluări
- C 10 - Acizii Nucleici ProteineDocument27 paginiC 10 - Acizii Nucleici ProteineSergiuR94Încă nu există evaluări
- C 11 - Conversia Energiei În Celule, Mecanismul Molecular Al Fosforilării Oxidative Mitocondriale.Document23 paginiC 11 - Conversia Energiei În Celule, Mecanismul Molecular Al Fosforilării Oxidative Mitocondriale.SergiuR94Încă nu există evaluări
- C 5 - Compartimente Intracelulare Și Transport. Sortarea Proteinelor, Transportul Vezicular, Căi de Secreție Celulară.Document38 paginiC 5 - Compartimente Intracelulare Și Transport. Sortarea Proteinelor, Transportul Vezicular, Căi de Secreție Celulară.SergiuR94Încă nu există evaluări
- Biofizica Analizatorului VizualDocument50 paginiBiofizica Analizatorului VizualSergiuR94Încă nu există evaluări
- C 6 - Semnalizarea CelularaDocument24 paginiC 6 - Semnalizarea CelularaSergiuR94Încă nu există evaluări
- Etapele Ce Preced Implantarea - Celule Sexuale - Spermatozoidul.Spermiogeneza - AnomaliiDocument23 paginiEtapele Ce Preced Implantarea - Celule Sexuale - Spermatozoidul.Spermiogeneza - AnomaliiSergiuR94Încă nu există evaluări
- Pag 2 Referat 1Document2 paginiPag 2 Referat 1SergiuR94Încă nu există evaluări
- Apa, Biofizica Sistemelor Disperse 4.Document59 paginiApa, Biofizica Sistemelor Disperse 4.SergiuR94Încă nu există evaluări
- C 3 - Membranele CelulareDocument56 paginiC 3 - Membranele CelulareSergiuR94Încă nu există evaluări
- C 2 - Organizarea Chimica A CeluleiDocument35 paginiC 2 - Organizarea Chimica A CeluleiSergiuR94Încă nu există evaluări
- Pag 2 Referat 2Document4 paginiPag 2 Referat 2SergiuR94Încă nu există evaluări
- Anatomie. Introducere. Obiect. Embriologia.Document31 paginiAnatomie. Introducere. Obiect. Embriologia.SergiuR94Încă nu există evaluări
- Fecundatia - Zigotul.Perioada de Crestere Si Dezvoltare - Stadiul de Morula - Stadiul de BlastulaDocument15 paginiFecundatia - Zigotul.Perioada de Crestere Si Dezvoltare - Stadiul de Morula - Stadiul de BlastulaSergiuR94100% (1)
- Caracterele Morfologice Si Functionale Ale Dezvoltarii in Perioada Fetala. Lunile 3-9 de Viata IntrauterinaDocument39 paginiCaracterele Morfologice Si Functionale Ale Dezvoltarii in Perioada Fetala. Lunile 3-9 de Viata IntrauterinaSergiuR94Încă nu există evaluări
- Pag 2 Referat 2Document6 paginiPag 2 Referat 2SergiuR94Încă nu există evaluări