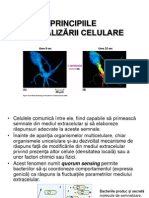
S-ar putea să vă placă și
- Structura GeneiDocument7 paginiStructura GeneiBianca OlteanuÎncă nu există evaluări
- Citogenetica MolecularaDocument13 paginiCitogenetica MolecularaAlexandra ViolettÎncă nu există evaluări
- Arms PCR 2Document4 paginiArms PCR 2Paul CheregiÎncă nu există evaluări
- Imunofenotipare LimfocitarăDocument15 paginiImunofenotipare LimfocitarăCristina Paun100% (1)
- Tehnici de BlottingDocument9 paginiTehnici de Blottingovidiu popaÎncă nu există evaluări
- Southern BlottingDocument10 paginiSouthern Blottingchris_ty_an7256Încă nu există evaluări
- Lab 1 - Principii Izolare ADNDocument55 paginiLab 1 - Principii Izolare ADNAntonia BadeaÎncă nu există evaluări
- Curs 5 Mutatii 14 OctDocument129 paginiCurs 5 Mutatii 14 OctCaraiman RalucaÎncă nu există evaluări
- Clase de Gene Implicate in Dezvoltarea CanceruluiDocument9 paginiClase de Gene Implicate in Dezvoltarea CanceruluiMihaela Țarnă100% (1)
- Curs 3Document83 paginiCurs 3Ramona UrzicaÎncă nu există evaluări
- ImunofluorescentaDocument1 paginăImunofluorescentaCucută Alexandru-DanielÎncă nu există evaluări
- AciziiNucleici 2018 Ambros1480018649Document157 paginiAciziiNucleici 2018 Ambros1480018649Luciana VornicÎncă nu există evaluări
- Suport Curs Genetica MedicalaDocument203 paginiSuport Curs Genetica MedicalaRaluca IoanaÎncă nu există evaluări
- CURS GU 2018 Partea I PDFDocument108 paginiCURS GU 2018 Partea I PDFCătălina VetrilaÎncă nu există evaluări
- MHC CursDocument31 paginiMHC CursBogdan AdrianaÎncă nu există evaluări
- Curs 2 - Metode Moleculare de Analiza A Acizilor NucleiciDocument37 paginiCurs 2 - Metode Moleculare de Analiza A Acizilor NucleiciPatricia Hera100% (1)
- Hipercolesterolemia FamilialaDocument8 paginiHipercolesterolemia FamilialaRamona ArseneÎncă nu există evaluări
- Genetică MedicalăDocument9 paginiGenetică MedicalăMarius LețÎncă nu există evaluări
- Ursu Ramona GabrielaDocument62 paginiUrsu Ramona Gabrielahefaistos2Încă nu există evaluări
- Imunofenotipare LimfocitaraDocument13 paginiImunofenotipare LimfocitaraRomeo-Remus RaffaiÎncă nu există evaluări
- Elemente Celulare Cu Rol Genetic 1Document13 paginiElemente Celulare Cu Rol Genetic 1Anda OajdeaÎncă nu există evaluări
- Curs GeneticaDocument42 paginiCurs GeneticaAlexandra MihaiÎncă nu există evaluări
- Curs 11. AntigeneleDocument6 paginiCurs 11. Antigenelejhonny cashÎncă nu există evaluări
- Grupe SanguineDocument12 paginiGrupe Sanguineveve1234Încă nu există evaluări
- Imuno-Cooperare CelularaDocument8 paginiImuno-Cooperare CelularaBianca BradÎncă nu există evaluări
- Metodele PCR Utilizate Pentru Deter Min A Rile de Biologie Molecular ADocument71 paginiMetodele PCR Utilizate Pentru Deter Min A Rile de Biologie Molecular AFlorea Rodica100% (1)
- Analiza CromozomilorDocument53 paginiAnaliza CromozomilorPatricia Beznea100% (2)
- Fisa Disciplinei Biologie CelularaDocument3 paginiFisa Disciplinei Biologie CelularaihamihaÎncă nu există evaluări
- Cromozomii 22Document18 paginiCromozomii 22Ama PredaÎncă nu există evaluări
- Imunologie Curs 2Document5 paginiImunologie Curs 2flori.bÎncă nu există evaluări
- Genetica Curs 1Document13 paginiGenetica Curs 1Alina Kacmaz100% (1)
- CELULADocument7 paginiCELULAMadalina FoteaÎncă nu există evaluări
- Prez EpigeneticaDocument23 paginiPrez EpigeneticaNeacsu OvidiuÎncă nu există evaluări
- Tehnica RFLP.Document17 paginiTehnica RFLP.Rodica Circea100% (1)
- Apoptoza Si Imunologia TumorilorDocument73 paginiApoptoza Si Imunologia TumorilorGeorge Mihail ManÎncă nu există evaluări
- Referat docxVIOLETADocument10 paginiReferat docxVIOLETAtuturici liviuÎncă nu există evaluări
- Imunol P I FinalDocument193 paginiImunol P I FinalMaria BadeaÎncă nu există evaluări
- Pirosecvențierea Și Tehnologia 454Document3 paginiPirosecvențierea Și Tehnologia 454Naomi TriteanÎncă nu există evaluări
- AQUAPORINEDocument26 paginiAQUAPORINEProca IonÎncă nu există evaluări
- Biologie Celulara Curs 2Document4 paginiBiologie Celulara Curs 2Voineta Gheorghiu100% (1)
- Lab 2 - Izolare ADN Bacterii Si PuritateDocument25 paginiLab 2 - Izolare ADN Bacterii Si PuritateAntonia BadeaÎncă nu există evaluări
- Curs 4 Genetica 20 Martie 2014Document60 paginiCurs 4 Genetica 20 Martie 2014Maria Cristina NaeÎncă nu există evaluări
- Izolarea (Extractia Adn Genomic Din Sange)Document3 paginiIzolarea (Extractia Adn Genomic Din Sange)George Dan IonÎncă nu există evaluări
- Prezentare BCHE ModifDocument499 paginiPrezentare BCHE ModifRoxana Nae PuccaÎncă nu există evaluări
- INGINERIAGENOMICADocument20 paginiINGINERIAGENOMICAMarius StancuÎncă nu există evaluări
- Metode Uzuale in Screenenigul Si DiagnosticulDocument272 paginiMetode Uzuale in Screenenigul Si DiagnosticulMada madalinaÎncă nu există evaluări
- Biologie MolecularaDocument62 paginiBiologie MolecularageorgemihailÎncă nu există evaluări
- Biologie CelularaDocument18 paginiBiologie CelularaRares IonutÎncă nu există evaluări
- Ps-00 Reactii de AglutinareDocument22 paginiPs-00 Reactii de AglutinareAndreea DeacÎncă nu există evaluări
- Anomalii Cromozomiale Asociate Cancerului UmanDocument2 paginiAnomalii Cromozomiale Asociate Cancerului UmanOana ElenaÎncă nu există evaluări
- SemnalizareaDocument82 paginiSemnalizareaAnna YepÎncă nu există evaluări
- Hierarchical Shotgun SequencingDocument2 paginiHierarchical Shotgun SequencingAndreea Elena BÎncă nu există evaluări
- Genele HousekeepingDocument13 paginiGenele HousekeepingMaria VdovicencoÎncă nu există evaluări
- Rezolvarea Subiectelor BDADocument17 paginiRezolvarea Subiectelor BDAAlex Cosmin100% (1)
- Profilaxia Bolilor GeneticeDocument13 paginiProfilaxia Bolilor GeneticeAndreea PrisacariuÎncă nu există evaluări
- Proprietăţile Și Funcţiile Sistemului HlaDocument6 paginiProprietăţile Și Funcţiile Sistemului HlaAndreea Mrsc50% (2)
- Curs 2Document83 paginiCurs 2Cristi AndreiÎncă nu există evaluări
- Curs I GeneticaDocument5 paginiCurs I GeneticaCristina TomaÎncă nu există evaluări
- Curs 2 Alimente Modificate Genetic-1Document23 paginiCurs 2 Alimente Modificate Genetic-1mihailmadalinosÎncă nu există evaluări
- Codul GeneticDocument7 paginiCodul GeneticAlina TeodoraÎncă nu există evaluări
- Hepatitele Acute Virale Induse A, B, C, D, E, G.Document31 paginiHepatitele Acute Virale Induse A, B, C, D, E, G.anastasia buzaÎncă nu există evaluări
- Tehnici de Biololie MolecularaDocument32 paginiTehnici de Biololie Molecularaanastasia buzaÎncă nu există evaluări
- Izolarea Si PurificareaDocument16 paginiIzolarea Si Purificareaanastasia buza100% (1)
- A.I. Iso 15189 - P1Document170 paginiA.I. Iso 15189 - P1anastasia buza100% (1)
- ElectroforezaDocument21 paginiElectroforezaanastasia buzaÎncă nu există evaluări
- Genetica Umana Lectia3Document40 paginiGenetica Umana Lectia3anastasia buzaÎncă nu există evaluări
- Prezentare OxidoreductazeDocument37 paginiPrezentare Oxidoreductazeanastasia buzaÎncă nu există evaluări
- Clonarea ADNDocument41 paginiClonarea ADNanastasia buzaÎncă nu există evaluări
- Genetica Umana Lectia4Document16 paginiGenetica Umana Lectia4anastasia buzaÎncă nu există evaluări
- Prelucrarea AutomatizataDocument20 paginiPrelucrarea AutomatizataVasea BickuÎncă nu există evaluări
- Genetica Umana Lectia2Document26 paginiGenetica Umana Lectia2anastasia buzaÎncă nu există evaluări
- Enzime 2Document56 paginiEnzime 2anastasia buzaÎncă nu există evaluări
- Reacția Autoimună - Sistemul ImunitarDocument22 paginiReacția Autoimună - Sistemul Imunitaranastasia buzaÎncă nu există evaluări
- Markerii Molecular-Genetici Si Epigenetici in PatologiileDocument14 paginiMarkerii Molecular-Genetici Si Epigenetici in Patologiileanastasia buzaÎncă nu există evaluări
- Ordin 1279Document12 paginiOrdin 1279anastasia buzaÎncă nu există evaluări
- Manual Utilizare IMMULITE 2000Document259 paginiManual Utilizare IMMULITE 2000Gabriela Ion Alexandrescu100% (1)
- 6 конвертированDocument16 pagini6 конвертированhellcatÎncă nu există evaluări
- Program - Calculul Incertitudinii de MăsurareDocument1 paginăProgram - Calculul Incertitudinii de Măsurareanastasia buzaÎncă nu există evaluări
- HG nr.201 Din 28.03.2017Document8 paginiHG nr.201 Din 28.03.2017anastasia buzaÎncă nu există evaluări
- Ghid Hemocultura Final 16.10.2020Document58 paginiGhid Hemocultura Final 16.10.2020NadejdaÎncă nu există evaluări
- HG 944 Din 03.10.2018 Regulament Etica MedicamentelorDocument7 paginiHG 944 Din 03.10.2018 Regulament Etica Medicamenteloranastasia buzaÎncă nu există evaluări
- HG 966 Din 18.10.2010 Regulament de Indicare PreturilorDocument7 paginiHG 966 Din 18.10.2010 Regulament de Indicare Preturiloranastasia buzaÎncă nu există evaluări
- Anexa 1 - Validarea MetodelorDocument11 paginiAnexa 1 - Validarea Metodeloranastasia buza100% (1)
- ANEXA 5, Program Pentru Compararea Metodelor Pe Baza Diagramei DiferenţelorDocument2 paginiANEXA 5, Program Pentru Compararea Metodelor Pe Baza Diagramei Diferenţeloranastasia buzaÎncă nu există evaluări
- Curs Cap 4 Compararea MetodelorDocument7 paginiCurs Cap 4 Compararea Metodeloranastasia buzaÎncă nu există evaluări
- ANEXA 4, Program Pentru Compararea Metodelor Pe Baza Testului T-PerecheDocument5 paginiANEXA 4, Program Pentru Compararea Metodelor Pe Baza Testului T-Perecheanastasia buzaÎncă nu există evaluări
- Curs Cap 3 Control de CalitateDocument13 paginiCurs Cap 3 Control de Calitateanastasia buzaÎncă nu există evaluări
- Curs Cap 2 Material de Referinta. TrasabilitateDocument8 paginiCurs Cap 2 Material de Referinta. Trasabilitateanastasia buzaÎncă nu există evaluări
- Curs CAP 1 Aspecte de Metrologie AplicataDocument14 paginiCurs CAP 1 Aspecte de Metrologie Aplicataanastasia buzaÎncă nu există evaluări