Documente Academic
Documente Profesional
Documente Cultură
Fiziologie Subiecte 2009 - Rezolvate
Fiziologie Subiecte 2009 - Rezolvate
Încărcat de
Merry LeeDrepturi de autor
Formate disponibile
Partajați acest document
Partajați sau inserați document
Vi se pare util acest document?
Este necorespunzător acest conținut?
Raportați acest documentDrepturi de autor:
Formate disponibile
Fiziologie Subiecte 2009 - Rezolvate
Fiziologie Subiecte 2009 - Rezolvate
Încărcat de
Merry LeeDrepturi de autor:
Formate disponibile
Fiziologie subiecte 2009
1.Functia de transport a sistemului cardiovascular
Debit cardiac 5 l /min. La 75 de ani=00 milioane l. !arametrii circulatori " presiune si #lu$ sanguin.
Distributia sangelui " %& in circulatia sistemica' 9 in circulatia pulmonara si 7 in inima.(emni#icatii
#unctionale " substante nutritive' metaboliti' subst cu rol de semnalizare' transport termic
)termoreglare*
Debitul cardiac este de 5l/min
(trec+ receptorii controleaza volumul de sange si se a#la in zone #oarte ,oase. La nivel coronar' debitul
este de 5&.
Debitul arterial ramane la o valoare constanta.
2.Functia de secretie " peptidul atrial natriuretic
!eptidul atrial natriuretic' -.)/D0F*1 #actor derivat din endoteliu ce produce vasodilatatie si
agregarea plac+etara' prostaciclina !232' si endotelina vasoconstrictor 4/5 si /462 ' vasodilatator
/461.Factori implicati in /F7 " ec+ilibrul #luido coagulant
!5- #avorizeaza eliminarea sodiului urina
Factorii implicati in ec+ilibrul #luidor coagulant /F7 " apare un c+eag de sange " in#arct. Dependent
de viteza unor substante la nivelul endoteliului. 3n piscina se stimuleaza intoarceea veneoasa.
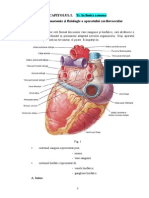
8.(istemul valvular al inimii
Valvele atrio-ventriculare -
localizate ntre atrii i ventriculi:
-valva mitral (M) - ntre AS i VS.
-valva tricuspid (T) - ntre AD i VD.
Valvele simoidiene ! localizate ntre ventriculi i marile artere:
-valva aortic (A) - ntre VS i aort.
-valva pulmonar (") - ntre VD i artera pulmonar.
Sistemul valvular :
#iecare valv are trei cuspe$ e%cept&nd valva mitral$ care are numai dou cuspe
Valvele previn reuritarea s&nelui.
Asiur curerea unidirec'ional$ esen'ial pentru (unc'ionarea inimii ca pomp )
curerea s&nelui numai din vene * atrii * ventriculi * aort sau artera
pulmonar$ dar nu i n sens invers.
Valvele cardiace pot prezenta dou tipuri de dis(unc'ii:
-insu+cien'a: valvele nu se mai nc,id complet$ determin&nd re-uarea
(reuritarea) s&nelui.
-stenoza: desc,iderea valvelor este redus sau se realizeaz reu. /nima tre0uie
s dezvolte o (or' mai mare pentru a mpine s&nele prin ori+ciul stenozat.
-valvele pot prezenta una sau am0ele tipuri de dis(unc'ii n acelai timp
(insu+cien'a i stenoza).
.!eptidul atrial natriuretic
"eptidul atrial natriuretic (A1").
- +0rele musculare din atrii au ranule asemntoare cu celulele landelor
endocrine care secret ,ormonii polipeptidici.
- A1" este secretat ca un prepro,ormon$ care se trans(orm ntr-un propeptid
stocat n ranule. Din acest propeptid se eli0ereaz A1" (23 AA) ca (orm
circulant
n s&ne.
- A1" se eli0ereaz n urma stimulrii receptorilor atriali de ctre creterea
volemiei$ a 1a4plasmatic$ n cazul e%cesului de aniotensin // circulant i la
creterea #5.
- alimentele cu pu'in 1a4in,i0 eli0erarea de A1"
6ezultatele ac'iunii A1":
6ezultat principal: 7 presiunii venoase centrale i a presarcinii.
8
9a nivel cardiac: relarea pe termen scurt a ,emodinamicii la creterile 0rute ale
volemiei.
5reterea A1" n ta,icardiile paro%istice e%plic natriureza care le nso'ete.
:n insu+cien'a cardiac conestiv$ A1" crete compensator pentru a reduce
ncrcarea inimii$ prin
creterea eliminrii renale de 1a4i ap
5./ndotelina
(ubstanta vasoconstrictoare produsa de endoteliu " #actorul constrictor derivat din endoteliu
este un puternic vasoconstrictor$ cu e(ect asemntor aniotensinei // i mai
puternic dec&t al neuropeptidului ;.
<D5# este implicat n modi+carea TA i n patoenia =TA$ aterosclerozei$
cardiopatiei isc,emice i a
insu+cien'ei renale si eli0erarea ei creste la (ri.
/ntervine in patoeneza aterosclerozei$ cardiopatiei isc,emice.
9.Distributia sangelui in sistemul cardiovascular
%&1circulatia sistemca' 9& circulatia pulmonara'7&inima' 9& vene.
La s#arsitul diastolei degenra in ventriculul drept1:1001120;volumul ventricular1:1litru
Debitul sanguin renal " 20125& din debitul cardiac' 1200ml/min. <nele #ibre a,ung la nivelul
+ipotalamusului " vasopresina' avand un e#ect de in+ibare " mecanism nervos.
7.3nervatia miocardului
/nerva'ia simpatic a inimii (nervii cardiaci)
8. >riinea (/ neuron): MS ! coarnele laterale T8-T?(3) 4@- ultimele dou 5$
2. +0re preanlionare (scurte) *sinapsa cu neuronul // n anlionii veetativi
simpatici cervicodorsali (n special anlionul stelat).
A. +0rele postanlionare (luni) ) Anervi cardiaci (superior$ miBlociu$ in(erior) )
(ormeaz ple% epicardice%tins.
C. inerveaz toate structurile cardiace
Ac'iunea S1VS asupra inimii predomin n condi'iile de
solicitare +zic i@sau psi,ic.
D. Elocarea F-receptorilor cu "ropranolol ) 7#5.
G. Elocarea selectiv a F8-receptorilor cu Atenolol ) 7#5.
8H. Elocarea concomitent a S1VS i S1V" duce la dispari'ia in-uen'elor
sistemului nervos autonom asupra inimii ) I #5 J 8HH 0ti@min (#5 intrinsec)$
datorat activit'ii intrinseci a 1S
/nervatia parasimpatica
>riinea (/ neuron): n 0ul0$ n nucleul dorsal al vaului (1DV) i nucleul am0iuu
(1cA) ) locul de plecare al nervilor vai.
2. +0re preanlionare Kluni * co0oarn mediastin *sinapsa neuronul // n din
anlionii veetativi parasimpatici$ localiza'i n peretele inimii.
A. +0rele post-anlionare K scurte.
C. inerveaz inima$ n special S<5 i Atriile
L. :n repaus$ asupra inimii predomin ac'iunea vaului.
D. Manevrele vaale (compresiunea lo0ilor oculari$
compresiunea sinusului carotidian$ manevra Valsalva -
e%pir cu lota nc,is) )IStimularea vaal )7#5
- Mtilitate:
<valuarea tonusului vaal.
>prirea unei ta,icardii paro%istice supra-
ventricular
%./#ectul stimularii parasimpatice asupra inimii
. <(ecte:
2
8) N-O "ropriet'i cardiace:
P cronotrop - ) 7 #5 (p&n la stop cardiac cu (enomen de scpare vaal)
P inotrop - ) 7 # contrac'ie
P tonotrop -
P dromotrop -
P 0atmotrop
Sec'ionarea nervilor vai sau 0locarea receptorilor
muscarinici cu Atropin (e(ect parasimpaticolitic
9./#ectul stimularii simpatice asupra inimii
10.7irculatia coronariana
8.Dou artere coronare$dinAo:art. coron. Dr *VD 4 VS "ost.
art. coron. St*VS Ant 4 9at.
2.Dispozi'ie: dinzona su0epicardic(artere mari cu IQ-6ec)*spre
zonasu0endocardic(artere mici i arteriole cu IF2-6ec).
A.Vene principale:sinus coronar(L?R s&ne)4vena cardiac ant.
(2HR s&ne) *AD
vene t,e0esiene*cavit'ile inimii
C.Tipuri de circula'ie:terminal.colateral
"er(uzia diastolica e mai mare decat cea sistolica.
#lu%ul sanuin coronarian in coronara dreapta e prezent atat in timpul sistolei
ventriculare cat si in timpul diastolei.
/n coronara stana e (oarte redus in timpul sistolei ventriculare in special in zona
su0endocardica$datorita compresiunii e%trinseci si este crescut in timpul diastolei
ventriculare
"resiunea sistolica e deDHmm= si cea diastolica de 2Hmm=
88.Sistemul e%cito conducator
<ste alcatuit din:
- nodul sinusal ce emite stimuli cu (recventa de 3H-8HH 0atai@min
- caile internodale atriale ! reprezinta cai pre(erentiale prin care se transmite
e%citatia la nivelul atriilor si nu sunt separate de +0re de colaen de restul
miocardului atrial
- nodul atrio ventricular are o (recventa de descarcare de CH-3H 0atai@min si
reprezinta sinura leatura electrica intre atrii si ventriculi. Toate structurile
sistemului e%cito conducator sunt situate deasupra1AV si sunt inervate vaal
- sistemul de conducere ventricular ! ramura comuna a (asciculului =is porneste
de la nivelul 1AV$ patrunde in septul +0ros$ continua la nivelul septului
intraventricular mem0ranos$ divizandu-se in doua ramuri: dreapta si stana.
6amura dreapta inerveaza ventriculul drept pana la peretele anterior si ape%ul$
iar ramura stana inerveaza (asciculul stan anterior ! peretele anterior al
ventriculului stan si musc,iul papilar anterior si (asciculul stan posterior
inerveaza musc,iul papilar posterior si restul peretelui ventriculului stan.
6eteaua "urSinBe reprezinta ramurile terminale ale (asciculului =is
18.!otentialul de actiune in miocard
Durata potentialului de actiune este de 800ms=0'8s )in #ibra striata potentialul de actiune este cel al
unui neuron=1ms*. !e durata potentialului de actiune miocardic se integreaza 2001800 potentiale de
actiune din #ibra sc+eletica.miocardul nu are contractie tetanica. Dupa 0'7 ms celula sc+eletica accepta
un alt stimul=perioada re#ractara inclusa in potentialul de actiune. La un stimul ce determina !5
canalele ionice sunt implicate si nu raspund la alt stimul. 3n celula miocardica perioada re#ractara
absoluta repr 75& din durata !5.3n #ibra striata #enomenul electric este de 100 ori mai mic decat cel
mecanic si nu e$ista #aza de platou.
15. !erioada re#ractara in miocard "
perioada in care tesutul nu raspunde la un stimul de intensitate normala.4ine cat potentialul de actiune.
!erioada re#ractara se imparte in doua perioade di#erite= perioada re#ractara absoluta " miocardul nu
raspunde la niciun stimul' orice intensitate ar avea acesta.7analele rapide de -a sunt desc+ise' se
A
inc+ide poarta de inactivare.!erioada re#ractara relativa " 50190mv " este #oarte scurta. 3n aceasta
perioada celula poate #i stimulata in anumite conditii )stimul de intensitate mult mai mare decat cea
normala' pot aparea potentiale de actiune*.!erioada +ipere$citabila tine din momentul atingerii
potentialului membranar de repaus pana se atinge ec+ilibrul ionic in celula.Daca actioneaza un stimul
mic' celula se poate totusi depolariza.3nima este ine$citabil> o lung> perioad> de timp 800
milisecunde' dac> un stimul ac?iunea asupra inimii' cardiomiocitul nu r>spunde deloc
indi#erent de intensitatea stimului = perioad> re#ractar> absolut>.La s#@rAitul platoului' doar un
stimul cu intensitate mare determin> un poten?ial' dar care nu se poate propaga. 5celaAi stimul
cu intensitate mare c>tre s#@rAitul pantei de polarizare Ai produce un poten?ial de ac?iune
propagabil
7u cat +iperpolarizarea este mai lunga celula nu raspunde la stimuli.0evenirea din +iperpolarizare la
perioada re#ractara se numeste repolarizare post+iperpolarizare si implica canale ionice dependente
care devin insensibile la potentialul electric.
3n celula miocardica perioada re#ractara absoluta =75& din durata potentialului de actiune' #enomenul
mecanic are o durata asemanatoare cu a celui electric.3n #ibra striata #enomenul eletcric este de 100 de
ori mai mic decat del mecanic nee$istand #aza de platou. Depolarizarea inimii incepe de pe #ata stg a
septului interventricular
19./$trasistola
7restrea calciului determina contractia. (B dureaza mai mult decat contractia unei #ibre = 0'8s'
diastola ocupa cea mai mare parte dintr1o secunda. !resiunea intraventriculara in diastola este
presiunea atmos#erica. 7ontractie prematura " pauza compensatorie " se sc+imba amplitudinea bataii.
Forta de contractie a ventriculului este direct proportionala cu lungimea #ibrei musculare de la s#arsitul
diastolei. Bolumul telediastolic este mai mare dupa e$trasistola.
17.7analele ionice ale celulei miocardice
(istemul de transport ionic membranar este #ormat din= canale ionice' pompe ionice si transportori
ionici.
7analele ionice " permit transportul con#orm gradientului de concentratie.
(unt de doua tipuri " controlate)gated* "de volta,' de mediator si mecanic " si canale #ara poarta )non
gated*1 transport de 1000 de ori mai lent decat in cele cu poarta.
7anale ionice de potasiu' de natriu rapide' de natriu si calciu' de natriu si clor. 4ransportori ionici pot #i
natriu si calciu' natriu +idrogen. !ompe ionice de natriu potasiu.
Cecanism de actiune= 7a2 " sarcolemale " comple$ 7a2 " calmodulina
1 sarcoplasmice " #os#olamban in+iba pompa
5analele de 1a4:n +0rele rapide (contractile): canale rapide de 1a4(voltaB-
dependente)$ desc,ise n (aza de depolarizare.n +0rele lente (pacemaSer):
canale speci+ce de 1a4(non-ated) sau (unnT c,annels$ activate n timpul D9D.
2. 5analele de U4(voltaB dependente i dependente de Ac,):desc,ise n (aza de
repolarizare.
rol principal: re(acerea poten'ialului de repaus i controlul nivelul e%cita0ilit'ii
celular.
Ac, le men'ine mai mult desc,ise$ induc&nd starea de ,iperpolarizare
21.7uplarea e$citatie contractie
Depolarizarea ventriculara dureaza 0'09s )mai putin decat timpul de conducere prin -5B pt ca -5B
are rol de temporizare a intrarii in actiune a ventriculului' sistola atriala alungeste sarcomerul. 7and
potetialul de actiune se propaga prin memnr transv se activeaza canalele de calciu ce duce la in#lu$ de
calciu in citoplasma.cuplarea dintre tubii 4 si reticulul sarcoplasmic este de natura mecanica. 7alciul
ce intra prin canalele din tubii 4 determina modi#icari spatiale ale canalelor din reticulul sarcoplasmic'
desc+izandu1le. !rin actiunea con,ugata a canalelor se declanseaza contractia. (c+imbatorul natriu
calciu e$clude calciu din celula in raport cu gradientul de concentratie al natriului care depinde de
pompa -a/D. /#ect inotrop pozitiv al digitalei " blocarea -a/D si 54! azei .0ela$area depinde de
7a.!ompa de calciu sarcolemala este controlata de calmodulina.!ompa de calciu din reticulul
sarcoplasmic este controlata de #o#solamban'calmodulina.
5ctiunea in+ibitorie a calmodulinei se mani#esta cand creste calciul in celula.
C
22.(istola ventriculara
1 sistola ventriculara care dureaza 0'80 s si se des#asoara in 2 #aze =
1 o #aza de punere in tensiune a #ibrelor musculare ventriculare' cand valvele atrioventriculare se
inc+id si ventriculele sunt pentru o perioada scurta cavitati inc+ise. 3n contiunare' presiunea creste
depasind1o pe cea din artere si ca urmare' se desc+id valvele semilunare si sangele este e,ectat in aorta
si artera pulmonara. 5ceasta #aza'care incepe in momentul inc+iderii valvelor atrioventriculare si se
termina cu desc+iderea valvelor semilunare' poarta numele de #aza de contractie izovolumetrica.
3n continuare' urmeaza #aza de e,ectie' cand sangele este e$pulzat in artera aorta si cea pulmonara.
Faza de e,ectie incepe cu desc+iderea valvelor semilunare si s#arseste cu inc+iderea acestora..
7antitatea de sange e,ectat de #iecare ventricul prin sistola ventriculara este de 70175 ml in repaus si
se numeste volum sistolic.
28.Diastola ventriculara
Dupa sistola urmeaza diastola ventriculara ce dureaza 0'50 s. 3n aceasta #aza' musculatura
ventriculara se rela$eaza' presiunea scade rapid si devine in#erioara celei din arterele mari
determinand inc+iderea valvelor acestora. 5st#el sangele nu re#lueaza inapoi in ventricule.
3ntre inceputul diastolei ventriculare si instalarea unei noi sistole atriale cavitatile inimii sunt rela$ate
)sunt in diastola*. 5ceasta #aza are durata de 0'0 s si se numeste diastola generala
2.Functia de pompa a inimii
Functia de pompa a inimii asigura deplasarea continua, intr-un singur sens a singelui.
Activitatea de pompa a inimii se desfasoara ciclic.
Descrierea miscarilor inimii ca intreg este di#icila daca se incearca aplicarea pentru intreg
cordul a modelelor #olosite pentru #ibra miocardica izolata' din mai multe motive=
1orientarea #ibrelor miocardice este comple$a' #ibrele ne#iind paralele ca la musc+iul papilar'
ci realizind o adevarata EbuclareF in interiorul peretilor inimiiG
1#ibrele musculare cardiace nu se contracta simultan' ci intr1o anumita secventa' dictata de
secventa de activare electrica a inimiiG
1cordul isi modi#ica #orma si grosimea in timpul contractieiG
1#orta e$ercitata in di#erite parti ale inimii este variabila in #unctie de curbura peretilor
cardiaci. B( are #orma unui cilindru cu un capat conoid'realizind aspectul unui elipsoid cu pereti
grosi. 3n timpul contractiei'el trece de la #orma elipsoidala la cea s#erica prin scurtarea a$ei vir#1
baza' datorata coboririi planului valvei
mitrale spre ape$' care urca spre baza inimii si prin alungirea a$ei transversale' asociata cu
ingrosarea peretilor ventriculari' ceea ce #ace ca per global' diametrul intracavitar transversal
sa scada. 5st#el' B( actioneaza ca o pompa de mare presiune' adaptata e$pulziei unor cantitati
importante de singe impotriva unor presiuni mari din aorta.
BD are #orma unei pungi' care #ace parte dintr1o s#era' septul 3B avind o #orma conve$a spre
cavitatea BD' iar peretele liber #iind asemanator unei calote s#erice care in#asoara septul.
/$pulzia singelui din BD se #ace prin trei mecanisme=
1scurtarea a$ului longitudinal prin contractia #asciculelor musculare si a mm papilari' ce trag
inelul valvei tricuspidei in ,osG
1contractia peretelui liber ventricular care este concav si se apropie ast#el de septul 3BG
1contractia #asciculelor miocardice circulare ale B(' care determina cresterea conve$itatii
septului 3B' micsorind a$ul transversal al BD. 5st#el' BD reuseste o miscare de Eburdu#F care
permite e$pulzia unei cantitati importante de singe #ara cresterea mare a presiunii
ventriculare. Di#erentele de geometrie intre cei 2 ventriculi permit pastrarea unui volum
sistolic egal' in conditiile in care ei lucreaza in regimuri presionale #oarte di#erite.
25.diagrama volum presiune a ventriculului stang
B( are #orma unui cilindru cu un capat conoid'
realizind aspectul unui elipsoid cu pereti grosi. 3n timpul contractiei'el trece de la #orma
elipsoidala la cea s#erica prin scurtarea a$ei vir#1baza' datorata coboririi planului valvei
mitrale spre ape$' care urca spre baza inimii si prin alungirea a$ei transversale' asociata cu
ingrosarea peretilor ventriculari' ceea ce #ace ca per global' diametrul intracavitar transversal
sa scada. 5st#el' B( actioneaza ca o pompa de mare presiune' adaptata e$pulziei unor cantitati
?
importante de singe impotriva unor presiuni mari din aorta. 5st#el' cresterea presiunii
intraventriculare duce la cresterea tensiuniiG+ipertro#ia
ventriculara )ingrosarea peretilor ventriculari* ca raspuns la o suprasolicitare mecanica )ca in
stenoza aortica* determina scaderea tensiunii in perete' permitind mentinerea presiunii de
per#uzie intr1un mod mai economicG dilatatia ventriculara )cresterea diametrului cavitatii
ventriculare* creste tensiunea la perete' determinind cresterea consumului miocardic de o2.
29. legea inimii
Dupa modul in care lungimea #ibrei miocardice este implicata in aceste mecanisme'
se discuta despre o autoreglare heterometrica si una homeometrica.
Autoreglarea heterometrica.Legea inimii Franck-Starling. Presarcina
(preincarcarea) reprezinta volumul de singe care umple ventriculul la s#irsitul
diastolei' adica B4D ventricular. Lungimea #ibrelor miocardice variaza in #unctie de
acest volum. (e poate spune ca ventriculul Eeste in#ormatF la s#irsitul #iecarei diastole
asupra sarcinii volumice pentru sistola ce urmeaza. (tarling a aratat in 191 pe
preparatul cord1pulmon de ciine ca inima in situ' dar izolata de orice in#luenta
nervoasa e$trinseca poate dezvolta o #orta de contractie proportionala cu alungirea
diastolica a #ibrelor sale. /l a constatat ca presiunea si volumul e,ectiei sistolice sint
proportionale cu volumul umplerii diastolice ventriculare' acest comportament
speci#ic reprezententind ELegea inimiiF. Legea inimii reprezinta asadar' un mecanism de
autoreglare intrinseca
heterometrica a activitatii cardiace. 5cest mecanism reprezinta o importanta
modalitate de adaptare' deoarece permite inimii normale sa isi creasca debitul bataie
prin cresterea presiunii de umplere ventriculare. La om' vir#ul curbei de per#ormanta
ventriculara corespunde unei presiuni telediastolice ventriculare de 12mmHg' mai
mare decit cea normala " 5mmHg in B( si 2'5mmHg in BD. Cecanismul este ilustrat
de o familie de cure Starling, care atesta dependenta B( nu doar de presiunea
telediastolica ventriculara' dar si de contractilitate, proprietate intrinseca a
miocardului. 3n momentul cresterii bruste a presiunii in aorta' B( nu are #orta necesara pentru un
debit bataie egal cu cel precedent' si va e,ecta o cantitate mai mica de singe. 5st#el'
B4( ventricular va creste' datorita scaderii F/G diastola urmatoare va asigura un
volum telediastolic crescut' umplerea diastolica realizindu1se cu un surplus de singe'
ramas din e,ectia anterioara' iar sistola consecutiva va asigura un volum bataie
crescut' egal cu cel e$istent inaintea cresterii presiunii aortice. Cecanismul de
autoreglare este tot +eterometric' datorat alungirii sarcomerului. 7resterea pe lunga
durata a postsarcinii are insa e#ecte negative asupra per#ormantei cardiace. 3mportanta practica a
mecanismului FrancI1(tarling este corelata cu adaptarea
deitului cardiac in radicardie, cind prin alungirea diastolei' creste umplerea
ventriculara' determinind cresterea volumului ataie si corelarea deitului cardiac
acelor ! ventriculi, care #unctioneaza ca un sistem de pompe asezate in serie intr1un
circuit inc+is' debitelelor trebuind mentinute riguros egale.
27.7arotidograma
"A#$%&D$'#A(A sau pulsul carotidian este inregistrarea variatiilor de volum
ale arterei carotide in timpul e,ectiei B(. 4e+nica de inregistrare este prin
pletismografie fotoelectrica sau prin plasarea de traductori mecanici in dreptul
a. carotide, la nivelul marginii interne a S"(. 5naliza mot#ologica a 7. distinge 2
#aze= fa)a sistolica, si cea diastolica. Fa)a sistolica incepe cu punctul e )/*' ce
corespunde desc+iderii sigmoidelor aortice' la debutul e,ectiei B(' care survine la
*,*+-*,*, sec dupa debutul J1. 7uprinde unda anacrota, sau de percutie' cu o
ascensiune rapida pina la un vir# notat !. !unctul ! este atins in *,-*-*,-! sec' ce
reprezinta timpul de ascensiuneG urmeaza o unda in platou' sau usor descendenta'
terminata printr1o rotun,ire notata 7 si o unda rapid descendenta' intrerupta de o
incizura notata cu 3 )3*' incizura dicrota. 5ceasta se datoreaza inc+iderii valvelor
3
sigmoide aortice si survine la 0'0210'08 sec dupa componenta 5 a J2. !ortiunea
descendenta a undei' incepind de la punctul ! pina la incizura dicrota 3 este unda
catacrota. <neori' aceasta unda poate avea un al doilea vir#' datorita rezistentei
vasculare,asa cum se intimpla la virstnici.
Fa)a diastolica cuprinde unda dicrota, de reascensiune, datorata ciocnirii singelui
de valvele sigmoide inc+ise' care are tendinta de re#lu$ spre B( in acest moment.
5naliza cronologica a pulsului carotidian permite calcularea urmatoarelor intervale=
timpul de semiascensiune reprezinta timpul necesar undei anacrote pentru a a,unge
la ,umatate din amplitudinea ma$ima. Durata normala este de *,*.-*,*+ secunde.
5cest timp se coreleaza bine cu gradientul de presiune transaortic creat de e,ectia B('
de aceea el este utilizat pentru evaluarea contractilitatii /S sau a severitatii
steno)elor aortice. (orfologia " depinde de vite)a de e0ectie a /S, de deitul
sistolic si de re)istenta circulatorie sistemica
!1.2ugulograma
23'3L$'#A(A este inregistrarea gra#ica a pulsului venos ,ugular' #iind o
re#lectare retrograda a variatiilor de presiune determinate de ciclul cardiac la nivelul
5D. 4e+nica de inregistrare se bazeaza pe utilizarea de captatoare speciale la nivelul
,ugularei drepte' ea #iind mecanograma cel mai greu de realizat. 5naliza mor#ologica
distinge urmatoarele unde=
1Ea4, unda po)itiva ce corespundei contractiei atriale5 apare la *,*6-*,-! sec dupa
deutul undei P5
-7c4 corespunde deutului sistolei /D si inchiderii tricuspidei5
-depresiunea 784 coincide cu e0ectia /D si se datorea)a trecerii singelui din
venele 0ugulare in AD, care se umple5
-unda 7v4 po)itiva, se datorea)a umplerii AD, cu usoara crestere a presiunii
intraatriale, in portiunea sa ascendenta, si deschiderii tricuspidei (virful undei v)
si inceputul golirii AD in /D in portiunea sa descendenta5
-depresiunea 9 se datorea)a umplerii rapide a /D, si 7suctiunii4 singelui din
atriu de catre /D5
-unda h marchea)a sfirsitul umplerii lente (diasta)is) al /D. . ,ugulograma
normala se caracterizeaza prin relatia a:c:v si $:K. Lugulograma este utila pentru
aprecierea cordului drept. 3n #ibrilatia atriala dispare unda a si unda $ este stearsa'
c+iar mai mica decit unda K. <nda a este accentuata in cazul unui obstacol
tricuspidian )stenoza tricuspidiana' tromboza de 5D*' al scaderii compliantei BD
)+ipertensiune pulmonara' stenoza pulmonara* sau in di#erite aritmii' in care creste
cantitatea de singe din atrii' prin urmare 5D nu se goleste bine in BD in timpul
sistolei atriale= ritm nodal' ta+icardie ventricularaG unda $' datorata golirii ,ugularelor
in 5D poate #i accentuata in D(5 )de#ectul septal atrial* si diminuata in insu#icienta
tricuspida si #ibrilatia atriala. <nda v este accentuata in insu#icienta tricuspida )34* si
D(5G depresiunea K are panta rapida in 34' pericardita constrictiva si insu#icienta BD
si panta lenta in stenoza tricuspidiana
29.5pe$ocardiograma
AP:;$"A#D&$'#A(A ("A#D&$'#A(A /S) este inregistrarea grafica a
viratiilor produse de miscarile virfului inimii in timpul "", corespun)ind
activitatii mecanice a /S. 5ceste vibratii au o #recventa scazuta si sint responsabile
de unele componente ale zgomotelor cardiace. A este echivalentul unei
fonocardiograme inregistrate in anda frecventelor foarte 0oase. 4e+nica de
inregistrare consta in asezarea unui micro#on piezoelectric la nivelul sp B ic lmc' loc
unde se palpeaza socul ape$ian. 5naliza mor#ologica permite identi#icarea unor
momente ale 77. 5naliza cronologica permite masurarea unor intervale de timpi
sistolici, cu valoare diagnostica egala cu cele otinute pe alte mecanograme si mai
ales a intervalelor diastolice. A este singura mecanograma ce permite
L
determinarea cu acuratete a acestora.
<nda 5 corespunde sistolei atriale' coincide cu J si incepe la 0'0%10'12 sec de la
debutul undei !G 5mplitudinea ei creste in stenoza mitrala si cind creste presiunea
telediastolica a B(= stenoza aortica' insu#icienta B(G ea dispare in F5.
3ncizura 7 se datoreaza debutului contractiei izovolumetrice a B(G intervalul 7/
reprezinta 73B a B(G <nda / este vir#ul sistolic' ce coincide cu desc+iderea valvei
aorticeG unda H )+umerus* coincide cu inc+iderea valvei aorticeG perioada /1H este
!/B(G unda . este vir#ul diastolic'ce coincide cu desc+iderea mitraleiG intervalul H1.
este perioada de rela$are izovolumetricaG intervalul .1 F reprezinta umplerea rapida
ventriculara' iar punctul F marc+eaza s#irsitul acesteiaG perioada .17 este perioada de
umplere ventriculara totala )rapida' lenta "diastazis1 si sistola atriala*. <nda F este
ampla in supraincarcarea diastolica a B( )insu#icienta mitrala' insu#icienta aortica* si
redusa ca amplitudine in stenoza mitrala. <nda F coincide cu J8.
80.Fazele sistolei ventriculare
.data cu inceputul contractiei ventriculare' presiunea
intracavitara incepe sa creasca si determina inc+iderea brusca a valvei mitrale )C3*.5cest
moment' considerat ca inceputul sistolei ventriculare stingi' se observa atit pe
ec+ocardiogra#ie cit si pe #onocardiograma' ca prima vibratie ampla a zgomotului 3 )J1 sau
(1*. .data cu inc+iderea mitralei' ventriculul devine o cavitate inc+isa. 7ontractia sa nu poate
modi#ica volumul sanguin' ci determina modi#icari ale #ormei cavitatii' cresterea in continuare
#oarte abrupta a presiunii intracavitare si a stressului parietal' adica a tensiunii parietale'
de#inita ca #orta e$ercitata pe unitatea de lungime)dKne/cm* atit la nivelul #ibrei miocardice
unice' cit si la nivelul peretului ventricular.Legea Laplace permite calculul tensiunii parietale
ventriculare din #ormula= 4 = ! $ r/2+' unde 4 este tensiunea parietala' ! este presiunea
intracavitara ventriculara' r este raza cavitatii ventriculare si + este grosimea peretelui
ventricular.
4ermenii de stress parietal si tensiune parietala nu sint propriuzis sinonimi' deoarece stressul
este #orta ce actioneaza pe unitatea de supra#ata' masurata in dKne/cm2. 7ontractia #ibrelor
miocardice genereaza de #apt stress.
5ceasta #aza dureaza apro$imativ 0'05secunde si reprezinta faza de contractie izovolumetrica
(izometrica) sau contractia ventriculara izocora. Desi are o durata scurta' ea are un rol
important in antrenamentul bataie cu bataie al miocardului. 7ind presiunea din B( depaseste
valoarea de %0mmHg' )valoarea presiunii singelui in aorta* iar cea din BD depaseste putin
valoarea de %mmHg )valoarea presiunii singelui in artera pulmonara*' valvele semilunare se
desc+id si incepe urmatoarea #aza' faza de contractie ventriculara izotonica, sau de ejectie.
5ceasta #aza cuprinde la rindul ei faza de ejectie rapida si faza de ejectie lenta.
Fa)a de e0ectie rapida incepe odata cu desc+iderea valvelor semilunare aortice si
pulmonare.Din acest moment' B( comunica larg cu aorta' #ormind o cavitate cu 2
compartimente' in care presiunile evolueaza concordant. 3n timpul e,ectiei rapide' circa 2/8
din cantitatea de singe din ventriculi sint e$pulzate cu viteza mare in aorta si
pulmonara./,ectia rapida cuprinde scurtul interval de timp dintre desc+iderea sigmoidelor si
atingerea valorii ma$ime a presiunii intraventriculare. 5ceste valori sint de -!*mm<g
pentru /S si !=mm<g pentru /D.Di#erentele presionale dintre cei doi ventriculi se
datoreaza #ortei de contractie di#erite ale celor doi ventriculi si compliantei mai mari a arterei
pulmonare. Desi in conditiile unei #recvente cardiace normale e,ectia rapida reprezinta in ,ur
de 1/8 din totalul #azei de e,ectie' in acest timp ea determina golirea a 90& din volumul
bataie.
:0ectia lenta este cuprinsa intre vir#ul presiunii ventriculare si momentul inc+iderii valvelor
semilunare. 3n aceasta #aza' presiunile din ventriculi si arterele mari scad' atit datorita
incetinirii contractiei ventriculare' cit si datorita golirii ventriculilor si acumularii singelui in
vasele mari.3naintea terminarii e,ectiei' datorita golirii ventriculilor si umplerii vaselor mari'
gradientul presional dintre acestea se inverseaza' dar singele continua sa curga inspre vasele
D
mari in intervalul denumit protodiastola' considerat interval de inertie.Comentul inc+iderii
valvelor semilunare este marcat pe curba de presiune din vasele mari de o mica incizura'
urmata de o unda usor ascendenta,incizura dicrota. 5ceasta se datoreaza tendintei singelui de
a se intoarce spre ventriculi in momentul inc+iderii valvelor sigmoide si izbirii de valvele
inc+ise' ce determina o discreta ascensiune presionala. !e parcursul e,ectiei' ventriculii se
golesc in special prin reducerea diametrului lor transversal' volumul e,ectat in vasele mari
#iind de apro$imativ 6*ml, denumit volum bataie sau volum sistolic.7antitatea de singe
ramasa in ventricul la s#irsitul e,ectiei se numeste volum telesistolic' valoarea sa normala
#iind de 50190ml. 0aportul dintre volumul sistolic si volumul telediastolic )de la s#irsitul
umplerii ventriculare* se numeste fractie de e0ectie. Durata e,ectiei rapide este de
*,*,secunde' iar a celei lente de *.->secunde.
81 Fazele diastolei ventriculare
Fa)ele diastolei ventriculare.(#irsitul protodiastolei #iziologice este marcat de inc+iderea
valvelor semilunare aortice si pulmonare. #ela8area i)ovolumetrica este cuprinsa intre
momentul inc+iderii valvelor semilunare si cel al desc+iderii valvelor atrio1ventriculare.3n
timpul acestei #aze' ventriculii sint cavitati inc+ise etans.
0ela$area izovolumetrica se termina atunci cind presiunea din ventriculi scade sub nivelul
presiunii din atrii' care in acest interval de timp creste progresiv' datorita intoarcerii venoase.
5cum se desc+id valvele atrio1ventriculare si incepe umplerea ventriculara.5ceasta #aza are o
durata de *,*1sec. !rocesul de rela$are este ca si cel contractil' un proces activ' care consuma
15& din energia consumata de miocard. /a necesita recaptarea 7a 2; #i$at de 4n17 de catre
0('eveniment ce permite disocierea actinei de miozina. 0ela$area izovolumetrica are loc in
timpul sau imediat dupa inlaturarea 7a activator din locurile de #i$are ale 4n17' prin actiunea
pompei de 7a de la nivelul 0(. 0ela$area izovolumetrica poate #i a#ectata atunci cind
transportul 7a in 0( intirzie' sau cind a#initatea 4n17 pentru 7a creste. 5ceste modi#icari
patologice pot #i cauzate de alterari structurale ale 0( sau de scaderea concentratiei de 54! in
miocard.
Fa)a de umplere ventriculara rapida incepe imediat dupa desc+iderea valvelor
atrioventriculare.(ingele patrunde cu viteza in ventriculi' pe baza gradientului presional
atrioventricular.
Biteza de curgere a singelui la nivelul ori#iciului mitral este de *,,m?sec. 3n timpul
acestei #aze' se realizeaza apro$.2/8 din umplerea ventriculara' dar ponderea acestei #aze
variaza in #unctie de #recventa cardiaca' de supra#ata ori#iciului mitral si de complianta
ventriculara.!rin complianta intelegem raportul dintre variatia de volum dB si variatia de
presiune' d!' adica dB/d!. BD este mai compliant comparativ cu B(' datorita peretilor mai
subtiri. Durata acestei #aze este de *,--secunde. 0ela$area ventriculara in timpul #azei de
umplere rapida este in mica masura activa' rolul cel mai important #iind ,ucat de reculul
elastic pasiv al peretelui miocardic. 5st#el' are loc o adevarata EsuctiuneF ventriculara' care
#avorizeaza umplerea ventriculara.!entru cordul drept' suctiunea ventriculara se inregistreaza
pe ,ugulograma' sub #orma undei negative EMF. 7omplianta poate scadea in +ipertro#iile
ventriculare )ingrosarea peretilor ventriculari*' ceea ce duce la cresterea presiunii diastolice.
Fa)a de umplere ventriculara lenta incepe atunci cind debitul singelui prin valvele
atrioventriculare
scade'datorita scaderii gradientului presional atrio1ventricular prin golirea atriilor
si umplerea ventriculilor cu singe. 3n timpul umplerii lente' denumita diasta)is' are loc
cresterea lenta a volumului ventricular' in timp ce la nivel atrial presiunile scad lent' atingind
aproape un platou. Faza de umplere rapida si diastazisul se realizeaza numai pe baza
gradientului presional atrio1ventricular. /le au contributie ma,ora la umplerea ventriculara'
ponderea sistolei atriale #iind mult mai mica. Datorita di#erentelor de regimuri presionale si de
complianta ale arterelor mari' nu e$ista o per#ecta concordanta intre #azele ciclului cardiac
drept si sting. 5st#el' mitrala se desc+ide dupa tricuspida si se inc+ide inaintea ei' iar valva
aortica se desc+ide dupa valva pulmonara si se inc+ide inaintea ei.5ceasta #aza are o durata de
G
*,-,secunde.
82. Jgomotul 1 1 @- are o #recventa de 8010Hz si o durata de 0'1210'15secG el debuteaza la 0'021
0'0
sec dupa unda NG se asculta cel mai bine in sp 81 parasternal sting. /l are trei
segmente= initial' un segment de #recvente ,oase' datorat contractiei ventriculare in
perioada de mula, cind structurile ventriculare sint puse sub tensiune si se intind
corda,ele tendinoaseG segmentul mi,lociu' principal' datorat inc+iderii mitralei' cea
mai importanta componenta a acestui segment' urmata de inc+iderea tricuspideiG al
treilea segment se datoreaza accelerarii bruste a coloanei de singe la debutul e,ectiei
ventriculare. (egmentul terminal al J1 se datoreaza vibratiilor mici ale peretilor
arterelor mari )componenta vasculara*. J1 este mai amplu la copii si adolescenti' cu
perete toracic mai subtire' in sd.+iperIinetic' )ta+icardie*' in stenoza mitrala' in
stenoza tricuspidaG este scazut in amplitudine la obezi' em#izematosi' revarsat
pleuropericardic'3C5. Dedublat in 60D. (cade in intensitate in bradicardie si 65B
de gr.3.
88.Jgomotul 2 1 @! are o #recventa de 50170Hz si o durata de 0'0%10'10sec'avind o tonalitate mai
inalta. Debuteaza la s#irsitul undei 4. (e asculta cel mai bine in sp.2 parasternal. 3n
geneza vibratiilor ce alcatuiesc J2 intra vibratia de rela$are a ventriculilor' in timpul
rela$arii active' inc+iderea valvelor semilunare aortica si pulmonara' precum si
vibratiile coloanei de singe ce are tendinta la recul spre ventriculi. Datorita
asincronismului inc+iderii valvelor aortica si pulmonara' e$ista un decala, al celor
doua componente variabil in timpul respiratiei= in inspir' componentele se percep
dedublat' #iind separate de un interval de peste 0'0 sec' datorita modi#icarii umplerii
ventriculare in timpul inspirului. Bentriculul drept se umple mai bine in inspir'
datorita aspiratiei toracice crescute' datorata vidului pleural' in timp ce B( se umple
mai putin' datorita usoarei scaderi a intoarcerii singelui in 5( prin BB. !ulmonare.
5st#el componenta 5 a J2 apare mai devreme' iar componenta !1 mai tirziu' ceea ce
constituie dedublarea #iziologica a J2. 3n e$pir' #enomenele se petrec invers si
dedublarea dispare. 3n conditiile in care se alungeste e,ectia BD' dedublarea este si
mai evidenta = 60D' stenoza pulmonara' sau cind se scurteaza e,ectia B(= insu#icienta
mitrala. Dedublarea poate #i inversa )parado$ala* cind se alungeste e,ectia B(' iar
componenta 5 a J2 survine dupa componenta != 60(' stenoza aortica' 3C5. 3n aceste
situatii' in inspir' prelungirea #iziologica a e,ectiei BD #ace ca componenta !'
intirziind' sa se apropie de componenta 5' si sa se indeparteze in e$pir.
3n cazul unui D(5'in care se egalizeaza debitele B( si BD in timpul e$pirului si
inspirului' apare o dedublare larga' #i$a.
8 " la #el ca la 88
85 " <nda ! " e$presia eIg a depolarizarii atriale./ste rotun,ita pt ca depolarizarea nu e #oarte
mare in miocardul atrial.0eprezinta unda de depolarizare a atriului drept cobinata cu cea a atriului
stang.durata este de 10secunde.
891<nda N " prima unda negativa a comple$ului de depolarizare ventriculara N0( ce precede
prima unda pozitiva a acestuia.Durata ei este de ma$im 0'08 secunde. <nda N " prima unda
negativa neprecedata de una pozitiva.De obicei= proiectia vectorului septal' durata ma$ima 0'08
secunde si amplitudine ma$ima O din 01ul urmator.-u este obligatorie.Larga si/sau adanca
semni#ica necroza=in#arct cronic. /$ceptie " unda N pozitionala din D 333. <neori vectorul
principal se indeparteaza de D333.
87 <nda 4 "proiectia vectorului de repolarizare ventriculara. <nda rotun,ita si obligatoriu
asimetrica. De aceeasi parte a comple$ului N0(. <nda mica ce nu depaseste amplitudinea de 8
mm. 3nalta' ascutita dar asimetrica " la tineri sub 80 de ani. 4 vegetativ " tonus vegetativ e$agerat
"ascutita in #oaie de cort = +iperIalemie. (imetrica si sau inversata =isc+emie miocardica
8H
debutul este di#icil de apreciat pentru ca este o unda rotun,ita cu panta ascendenta de inclinatie
mai redusa.Debutul nu poate #i identi#icat cu usurinta pentru ca vectorii cresc in amplitudine si se
reinitiaza.0otun,ita asimetrica.!anta descendenta este mai abupta.
8%7omple$ul N0( "int oate derivatiile ma$im 0'0% s)#rontale*. 3n derivatiile precordiale 0'10.
7reste in +ipertro#ie ventriculara si in blocuri de ramura .!redomina 0 cu e$ceptia
B1'B2./c+idi#azic in B8.
<nda N " prima unda negativa neprecedata de una pozitiva.De obicei= proiectia vectorului septal'
durata ma$ima 0'08 secunde si amplitudine ma$ima O din 01ul urmator.-u este obligatorie.Larga
si/sau adanca semni#ica necroza=in#arct cronic. /$ceptie " unda N pozitionala din D 333. <neori
vectorul principal se indeparteaza de D333.
<nda 0 " obligatorie in toate derivatiile 0'0 s.dominanta cu e$ceptia B1'B2.3n mod
normal=proiectia vectorului principal de depolarizare B1'B2. /ste unda pozitiva a comple$ului
precedata sau nu de unda negativa.
<nda ( " negativa care urmeaza dupa o unda pozitiva. De obicei proiectia vectorului de baza
e$ceptie B1'B2. 5re 0'02 si nu este obligatorie.
unda P este prima unda negativa a comple$ului de depolarizare ventriculara N0( ce precede
prima unda pozitiva a acestuia. <nda 0 este prima unda pozitiva a comple$ului N0( si unda (
este unda negativa ce succede prima unda pozitiva a comple$ului adica unda 0.<nda N are o
amplitudine ma$ima 1/8 din unda 0 care urmeaza.
13ntervalul !N " durata semni#icatie.
(e masoara de la inceputul undei !' inceputul comple$ului ventricular. Dureaza 0'1210'20 s.
7riteriu important pt diagnostic de ritm sinusal. 7reste in blocuri 51B si necaracteristic in
+iperIalemii. (e scurteaza in sindrom de pree$citatie Q!Q.
2.Diagnostic de ritm sinusal
0itm sinusal=ritm normal .
<nda ! =depolarizare atriala " sensul normal al depolarizarii apare pe cel putin o
derivatie.3ntervale !N normale si constante.Biteza cardiaca intre 90190 batai/min.3ntervale 00
constante.7and undele ! nu au relatie cu N0(1 bloc 51B.3n ta+icardie si bradicardie se respecta
ritmul sinusal normal.
" originea in partea in#erioara a 5D in apropierea sinusului coronar.<nda ! apare inversata cu
mor#ologie normala " vectorul atrial se proiecteaza de ,os in sus. Qandering !acemaIer " in atrii
#unctioneaza pe rand mai multe #ocare ast#el incat " mor#ologie variabila a undelor ! si intervale
!N normale' dar variabile.
4a+icardie atriala multi#ocala " cauze " boala pulmonare severe ce a#ecteaza incarcarea 5D si BD
" +ipo$ie. 3ntra in #unctie #ocare ssupraventriculare ' undele ! au aspecte variabile si in unele zone
!N se poate scurta.
4a+icardie atriala paro$istica " un singur #ocar de depolarizare supraventriculara . in zona sup a
-5B . 1901220 batai/min. Cor#ologie unitara a undei ! " inversata in derivatii principale. R6loc
unidirectional cu reintrare= in sectiune transversala stimulul intra in #ibra musc' merge cu viteza
normala pe bratul sanatos si se opreste pe cel blocat. 7el de pe bratul sanatos nu se anuleaza )ca
in caz normal*' reintra pe bratul sanatos si descrie o miscare circulara ' consecinta = ta+iartmie
)incepe si se termina brusc*.
Flutter atrial= ta+iaritmie supraventriculara. 6loc unidirectional cu reintrare./D2 " lipsa liniei
izoelectrice )vectori multiplii o masc+eaza*' unde F ca valurile marii' intervale 00 variabile )unii
stimuli atriali cad pe -5B in perioada re#ractara si nu se mai transmit*.
Fibrilatie atriala = ta+iaritmie supraventriculara determinata de descarcare rapide de multiple
#ocare atriale mici = apar multi vectori la moment in secventa' nu e$ista unda ! pe traseu'
intervale 00 variabile.
0itm ,onctional " in ,onctiunea dintre -5B si Hiss. 3mpulsul spre atrii intarzie si este inversat' !
dupa N0( si inversata.
0itm de pace maIer " pace maIer arti#icial apare un spiIe urmeaza depolarizareventriculara'
comple$e N0( largite si inversate.
88
CA. 5alcularea a%ei electrice
A%ul electric se determina pentru ! atrii "$ ventriculi >6S si repolarizare
ventriculara T.
A%ul pentru V6S ! Metoda <int,oven: se ale 2 derivatii 0ipolare suma ale0rica
pt W$ r$ s $ se proiecteaza vectorul rezultant perpendicular din var(ul vectorilor
pana la intersectie- a%ul eletric e%primat ca orientare (ata de o orizontal$
manitudine$sens.
=e%ada ! nu se compara 0ipolara cu unipolara. 5and se (ace proiectia se tine
cont de X pozitiva a derivatiei. D / pozitiv la H rade$ D // la 3H$ D /// la 82H$ aV#
pozitiv la GH$ aV9 pozitiv la AH$ aV6 la 8?H.
Aspectele converent si diverent ies din cadranele normale.
C3. Depolarizarea atriala
Vectorul septal este neativ$ vectorul principal e pozitiv si vectorul de 0aza
neativ nu este o0liatoriu.
D// ! vectorul atrial pozitiv$ vectorul septal neativ$ vectorul principal poitiv si
mare$ vectorul de 0aza neativ si mic.
D/// ! vectorul atrial amplitudine mica pozitiv$ cel septal lipsa@neativ$ vectorul
principal este mai mic$ vectorul de 0aza este neativ.
6epolarizarea atriala de o0icei nu apare.
CD. Depolarizarea ventriculara
Vector principal de la dreapta la stana$ de sus in Bos$ dinspre anterior spre
posterior.VD se depolarizeaza dupa H$8G secunde$ iar VS dupa H$22s.Vectorul de
0aza de Bos in sus$ de la stana la dreapta. De la endocard la epicard ! comple%e
predominant pozitive.
CG. repolarizarea ventriculara
De la epi la endocard ! vectori de acelasi sens ! reteaua purSinBe- perioada
re(ractara (oarte luna$ potentialul de actiune dureaza AHHms.
?H."roteinele contractile ale miocardului
Miozina este principalul element al filamentelor groase. Ea are 2componente, denumite
meromiozine: meromiozina usoara si meromiozina grea.
Meromiozina usoara (light meromyosine LMM) este situata in portiunea centrala a
miofilamentului.Ea se agrega cu celelalte lanturi de meromiozina, pentru a forma filamentul
gros.
Meromiozina grea (heavy meromyosine HMM) formeaza capatul moleculei de miozina,
care se detaseaza de filamentul gros, formind punti transversale cu actina. Meromiozina
grea este formata din 2 subfragmente:
-subfragmentul 2 (S2) formeaza bratul moleculei de miozina; structura sa este
asemanatoare meromiozinei usoare
subfragmentul 1 (S1) formeaza capul moleculei de miozina, cu forma globulara; la
acest nivel sint localizate proprietatile fundamentale ale acesteia: activitatea ATP-azica
si capacitatea de a intereactiona cu actina.
Ca structura polipeptidica, molecula de miozina contine 2 lanturi grele (GM 200000D) si 4
lanturi usoare (GM 20000D). Cea mai mare parte a moleculei de miozina este formata din
cele 2 lanturi grele, spiralate in alfa-helix.
Lanturile grele ale miozinei sint de tip alfa si beta; lanturile alfa au viteza ATP-azica mai
mare. Dupa tipul de combinare a celor doua tipuri de lanturi, exista 3 izoenzime miozinice,
diferite ca viteza a activitatii ATP-azice:
-izoenzima V1, formata din 2 lanturi alfa;
-izoenzima V2, ce contine un lant alfa si unul beta;
-izoenzima V3, ce contine 2 lanturi beta.
La om exista un amestec aproximativ egal intre cele 3 izoenzime. In cazuri patologice, este
sintetizata preferential una sau alta dintre izoenzime. In hipertrofia ventriculara, se
sintetizeaza preferntial izoenzima V3, ceea ce permite o contractie mai economica. Acesta
este un adevarat mecanism de adaptare moleculara la anumite situatii patologice.
Cele 2 regiuni de flexibilitate ale lantului polipeptidic din structura miozinei se gasesc la zona
de separare dintre MM grea si usoara. Ele sint esentiale pentru modificarea unghiului dintre
82
componentele miozinei, fapt ce detine rol major in scurtarea fibrei miocardice.
Actina este principalul component al filamentelor subtiri. Ea are un capat fixat la nivelul
membranei Z, in timp ce celalalt aluneca printre filamentele de miozina. Actina exista in 2
forme;
Actina (G) globulara, monomer stabil, si actina F (fibrilara), forma polimerica,
caracteristica in vivo. Actina fibrilara este formata din 2 lanturi de monomeri care se
rasucesc in helix. Punctele de inflexiune sint plasate pe distanta a 7 monomeri de actina, la
intervale de 35-40 nm
?8."roteinele relatoare ale miocardului
Tropomiozina este o molecula in forma de bastonas, cu grosime de 2-3 nm.E formata din 2
lanturi polipeptidice, situate in jgheabul filamentului de F actina. Are o structura dublu-
helicoidala. Lungimea bastonasului de tropomiozina corespunde unei bucle a helixului de F-
actina.
Troponina se situeaza la punctele de inflexiune ale filamentelor subtiri de actina. Ea are 3
componente:
-troponina C, care leaga cu mare afinitate Ca,imediat ce concentratia acestuia in
sarcoplasma creste peste 10
-7
mol/L;
-troponina T, care leaga troponina de tropomiozina;
-troponina I care inhiba activitatea ATP-azica a miozinei, inhibind astfel interactiunea
dintre actina si miozina.
Rolul complexului troponina/tropomiozina este de a realiza controlul interactiunii actina-
miozina, deci al contractiei miocardice.
In starea de relaxare, la o concentratie scazuta a Ca intracelular, troponina C nu fixeaza Ca.
Ca urmare, tropomiozina ocupa intre filamentele de actina o pozitie de blocare, ce impiedica
steric interactiunea actina-miozina.
In timpul contractiei, declansata prin cresterea Ca intracelular, prin legarea acestuia de
troponina C se produce o schimbare a conformatiei moleculare. Astfel, se deplaseaza
tropomiozina din pozitia initiala, noua pozitie facind posibila interactiunea actina-miozina,
datorita expunerii locului de fixare a miozinei de pe suprafata G-actinei
Se poate spune despre Ca ca in mecanismul contractiei, el intervine prin impiedicarea
mecanismelor ce nu permit interactiunea actina-miozina: Ca are functie derepresoare
pentru contractilitate
?2.Mecanismul contractiei miocardice
Troponina si tropomiozina sint responsabile de reglarea contractiei miocardice, in
functie de disponibilul de ca intracelular.
Pentru mecanismul propriuzis al scurtarii fibrelor miocardice, este acceptata in continuare
teoria puntilor transversale, propusa de Huxley in 1957, care sustine formarea si desfacerea
repetitiv-ciclica a unor punti transversale intre filamentele subtiri de actina si cele groase de
miozina, realizate prin intermediul subfragmentului S1 al MM grele.
Secventa de realizare a acestor fenomene este:
-capul S1 al MM care contine fixat ADP si fosfat anorganic poate interactiona cu o actina
(globulara, monomer)prin legaturi electrostatice; in repaos, situsul de fixare de pe actina este
mascat de tropomiozina. Fixarea Ca la nivelul Troponinei C duce la expunerea acestui situs de
pe actina globulara, pe care se fixeaza fragmentul S1 al MM, ceea ce duce la schimbarea
unghiului dintre capul si gitul moleculei de miozina. Ca urmare, actina este trasa spre
mijlocul sarcomerului, ceea ce reprezinta mecanismul glisantal contractiei.
-formarea legaturii actina-miozina este urmata de eliberarea ADP si a fosfatului anorganic de
pe capul miozinic. Pentru desfacerea legaturii actina-miozina, este necesara prezenta de ATP
fixat pe capul S1 al miozinei. Cind ATP nu este disponibil, (ischemia miocardica), nu se
realizeaza desfacerea legaturii si apare contractura ischemica. Cind ATP este disponibil, el
se fixeaza pe subfragmentul S1, care se desprinde de actina. Datorita activitatii ATP-azice a
subfragmentului S1, ATP este scindat in ADPsi fosfat anorganic.
-fenomenul continua atita timp cit concentratia Ca la nivelul sarcomerului ramine ridicata.
Complexul troponina-tropomiozina permite realizarea interactiunii actina-miozina. Prin
repetarea succesiva a formarii si desfacerii puntilor transversale, filamentele subtiri aluneca in
8A
interiorul celor intunecate, cu scurtarea consecutiva sarcomerului
Forta de contractie depinde de Ca disponibil, iar viteza de contractie depinde de
activitatea ATP-azica a subfragmentului S1 al MM grele. Interrelatia este insa mult mai
complexa, cresterea cocentratiei Ca crescind de 5 ori aproximativ activitatea ATP-azica a
miozinei
?A.Distri0utia calciului la nivelul +0rei miocardice
Distributia Ca la nivelul fibrei miocardice
Studiul distributiei Ca se face prin tehnici radioizotopice, de histochimie si de microscopie
electronica, care nu pot insa diferentia Ca liber, ionizat, activ biologic, de formele neionizate.
In acest scop au fost utilizate tehnici speciale, cum este cea a microelectrozilor Ca-selectivi.
Datorita acestot tehnici, s-a demonstrat ca in miocit, Ca se afla in sarcolema, in RS,
mitocondrii, sarcoplasma si sarcomer. Au fost analizate si secventele dinamicii Ca in timpul
unui ciclu cardiac
Sarcolema indeplineste o serie de functii legate de dinamica ionului de Ca: mentine diferenta
de concentratie a acestuia la celula in repaos intre mediul extra si intracelular de
peste 10000 ori; permite schimburile rapide intre cele 2 medii pentru realizarea contractiei si
relaxarii si stocheaza cantitati importante de Ca la nivelul glicocalixului si glicoproteinelor
din structura sa. Schimburile sarcolemale de Ca se realizeaza prin intermediul canalelor
sarcolemale de Ca, al pompei sarcolemale de Ca si al antiportului Na-Ca
Aceste canale pot fi modulate sub actiunea neuromodulatorilor si altor cationi divalenti,
ca Mg. Catecolaminele stimuleaza activitatea acestor canale prin fosforilarea AMPc
dependenta a subunitatii alfa 1 din structura lor.
Farmacologic, canalele pot fi modulate prin blocantii canalelor de Ca, care se fixeaza pe un
situs receptor situat tot la nivelul subunitatii alfa 1. Exista minimum 3 clase distincte de
blocanti ai canalelor de Ca (dihidropiridinele-Nifedipina, fenilalkilaminele-Verapamilul,
benzotiodiazepinele-Diltiazemul), fiecare avind receptor specific astfel incit ele nu
competitioneaza pentru legarea de canal, ci se potenteaza reciproc. Este cunoscut si un
compus cu efect agonist al canalelor de Ca, care creste influxul acestuia prin actiune asupra
receptorului dihidropiridinic
Antiportul Na-Ca este asigurat de o proteina sarcolemala decelabila pe toata suprafata
sarcolemala, dar cu densitate maxima la nivelul tubilor T. Este principalul mecanism de
expulzare a Ca din miocit. Pentru 3Na introdusi in celula, in sensul gradientului de
concentratie, ea expulzeaza un Ca impotriva acestui gradient. Acest mecanism este indirect
controlat de ATP-aza Na-K dependenta, care mentine Na intracelular scazut intracelular,
necesar pentru intrarea acestuia la schimb cu Ca. Glicozizii digitalici blocheaza ATP-aza Na-
K dependenta, ceea ce favorizeaza incarcarea cu Ca a miocitului
?C.Modularea cuplului electro-contractil
Reprezinta secventa fenomenelor prin care se realizeaza legatura dintre activarea electrica a
celulelor miocardice si contractia lor.
Rolul esential revine Ca, ce mediaza interactiunea actina-miozina prin intrmediul sistemului
troponina-tropomiozina
Catecolaminele actioneaza prin stimularea receptorilor beta 1-adrenergici. Actioneaza
intracelular prin cresterea AMPc, datorita stimularii adenilatciclazei. Ca urmare, are loc
activarea proteinkinazei A, care va fosforila enzime intracelulare implicate in cuplul electro-
contractil:
-fosforilarea canalelor L sarcolemale va creste influxul Ca;
-fosforilarea dependenta de fosfolamban a pompei de Ca din RS favorizeaza captarea Ca;
-fosforilarea pompei sarcolemale de Ca favorizeaza expulzia Ca din celula;
-fosforilarea ATP-azei Na-K dependente favorizeaza expulzia Ca din celula, prin activarea
antiportului Na-Ca.
Stimularea receptorilor alfa 1 adrenergici activeaza fosfolipaza C, cu generarea
intracelulara de IP3, ce creste efluxul de Ca din RS prin intermediul receptorului pentru
IP3. Mecanismul este important pentru musculatura neteda vasculara, fiind incomplet
precizata importanta sa pentru miocit.
Ca urmare, catecolaminele favorizeaza contractia (efect inotrop pozitiv) dar si relaxarea (efect
lusitrop). Dezavantajul este cresterea consumului de O2.
Alti compusi actioneaza fie prin stimularea directa a adenilatciclazei, fie prin inhibarea
fosfodiesterazei, enzima implicata in degradarea AMPc. Toti determina, insa,cresterea
consumului miocardic de O2
8C
Glicozizii digitalici (Ouabaina, Digitala) blocheaza ATP-aza Na-K dependenta, fixindu-se pe
un receptor specific situat pe subunitatea alfa a acesteia. Se acumuleaza Na intracelular, ceea
ce reduce activitatea antiportului Na-Ca. Se modifica echilibrul dintre expulzia Ca din celula
si recaptarea sa in RS in favoarea recaptarii, ceea ce creste disponibilitatea Ca si deci forta de
contractie
Semnalul declansator al procesului contractil este declansarea potentialului de actiune.
(PA). Schimbarea brusca a potentialului membranar prin patrunderea Na in celula determina
deschiderea canalelor lente de Ca pe parcursul platoului potentialului de actiune
Influxul celular de Ca prin canalele lente nu modifica semnificativ concentratia intracelulara
a Ca, ci constituie un semnal (trigger) pentru eliberarea Ca stocat in mari cantitati in RS. Prin
tubii T, PA este condus in interiorul miocitului, in vecinatatea portiunii jonctionale a RS.
Cresterea concentratiei de Ca la acest nivel duce la activarea canalelor de Ca ryanodino-
senzitive din membrana RS, ceea ce duce la cresterea intracelulara a Ca datorita fenomenului eliberare de Ca
indusa de Ca. Ca se fixeaza pe troponina C, declansind contractia.
Cantitatea de Ca eliberata din RS depinde de cantitatea trigger de Ca ce intra prin canalele
lente L sarcolemale. Ca eliberat din RS nu poate activa canalele ryanodino-senzitive adiacente
care nu au fost activate de Ca patruns prin canalele L. RS poate elibera spontan mici cantitati
de Ca (Ca sparks), care nu declanseaza contractia, pentru ca nici Ca astfel eliberat nu
activeaza canalele adiacente ryanodino-senzitive. Atunci cind RS este supraincarcat cu Ca,
eliberarea spontana de Ca poate activa canalele adiacente, producind o adevarata unda
propagata ce creste semnificativ concentratia Ca in citosol, putind induce contractia. Astfel se
pot produce aritmii cardiace
55.Estimarea functiei diastolice a ventriculului
Este estimata prin masurarea timpului de relaxare izovolumetrica, de la inchiderea
valvelor sigmoide pina la deschiderea valvelor atrio-ventriculare (echocardiografic sau
mecanografic- apexocardiograma) si prin evaluarea umplerii ventriculare, pe baza relatiei
volum-presiune diastolica in timpul umplerii ventriculare, pe diagrama de lucru a VS. Panta
acestei curbe (dP/dV) reprezinta rigiditatea ventriculara, (stiffness ventricular) iar inversul
acesteia (dV/dP) este complianta ventriculara. Uzual, rigiditatea ventriculara se determina
pe parcursul diastazei
?3. proprietatile arterelor
Arterele sunt vase ce transporta su0 presiune sanele ce pleaca de la inima.
Arteriolele sunt valve de control ce eli0ereaza sane in capilare. Venulele
colecteaza sane din capilare si venele sunt conducte de transport al sanelui
spre inima$ rezervor maBor de sane$ repr 2@A din cantitatea de sane.cea mai
mare rezistenta este la nivelul arteriolelor.
<lasticitatea este capacitatea arterelor de a se destinde si a reveni la (orma
initiala in (unctie de variatiile de volum si presiune ale coloanei de sane din
interiorul lor. Trans(orma curerea sacadata in curere continua cu de0it crescut.
/n ,emoraii la pierderi de 2HR se mo0ilizeaza cantitatea de sane din sist
venos.
Alte proprietati:distensi0ilitate$ complianta$ contractilitate. 5ontractilitate ! prin
modi+carea cali0rului $ modi+carea rezistentei vasculare$ modi+carea de0itului
local in arteriole. <ste proprietatea arterelor de a-si sc,im0a cali0rul prin
contractia musculaturii netede. <ste ma%ima in arteriole. > presiune de per(uzie
adecvata tesutului si indepartarea cata0olitilor. S+ncterul arteriolar este un
ro0inet de presiune si 0araB pt socul sistolic. Dincolo de ele nu e%ista puls.
?L. "arametrii circulatiei arteriale
-de0itul arterial$ viteza de circulatie a sanelui$ circulatia laminara si tur0ulenta$
rezistenta vasculara$ presiunea arteriala$ lunimea vasului
De0itul arterial este determinat de rezistenta vascularaKimpedanta la -u% in acel
vas
Metode de masurare ! de0it metrul ultrasonic Doppler ! cristal piezoelectric
aplicat la supra(ata vasului. /n momentul activarii de catre un dispozitiv transmite
vi0ratii sonore coloanei de sane. > part a sunetelor sunt re-ectate de eritrocite
8?
ast(el incat undele sonore se intorc de la sane inapoi la cristal. Mndele re-ectate
sunt mai Boase decat cele incidente. Mnda re-ectata este reprimita si ampli+cata.
Viteza de circulatie a sanelui este dirept proportionala cu depitul si invers
proportionala cu sectiunea. 9eea lui Eernoulli: cu cat creste viteza de curere cu
atat scade presiunea laterale ce destinde vasul. /ntr-un vas cu placa de aterom
creste viteza de deplasare$ scade presiunea de distensie si permite
permanentizarea o0structiei.viteza scade in capilare si creste in venele cave.
5urerea laminaraKcurerea in straturi. C$L la 0ar0ati$ C$A la (emei si 8$D3 in
plasma. "roprietatile unui lic,id de a si sc,im0a treptat (orma datoritara (recarii
dintre elementele +urate. /mportanta in curerea sanelui: din intermitenta
devine continua si in rezistenta. 6ezistenta la curere a sanelui este de A ori mai
mare ca a apei si este nevoie de o presiune de A ori mai mare de impinere. <%:
anemii: viteza de circulatie a sanelui creste pt ca crete si (recventa cardiaca si
apare ta,icardia ! suprasolicitarea inimii.
5urerea tur0ulenta ! cand viteza de circulatie a sanelui creste$ la nivelul
supra(etei ruoase$ stenoze$cuduri$ depasind un pra critic se trans(orma in
curere tur0ulenta: sanele cure perpendicular pe vas in loc sa cura de-a
lunul luiK varteBuri care reduc viteza sanuina ! su-uriKmani(estare
stetacustica. Tipul de curere poate + apreciat prin numarul lui 6eTnolds cu
valoare normala su0 8HH. /n e(ort +zic viteza sanelui creste.
6ezistenta vasculara ! di+cultatea la curere intampinata de orice -uid ast(el
curerea se (ace cu consum de enerie$ iar plata se (ace in unitati de presiune
consumate pe parcurs. 5ea peri(erica se calculeaza$ nu se poate masura. Se
e%prima in dTne sec@cm sau unitati de rezistenta peri(erica (M6").K opozitia la
curere a unui lic,id a-at su0 radient de presiune de 8mm =. 8urp in
,ipertensiune pulmonara.
6ezistenta arteriolara ! la acest nivel rezistenta e ma%ima.
6T"- rezistenta totala peri(ericaK83HH-8LHH dTne
6 aortei si art mariK 8?H dTne
6 arteriolaraK8HHH dTne
6 capilaraK2HH dTne
6 venoasaK8?H dTne. 9a nivel de vas individual rezistenta art e mai mica decat
rezistenta cap. Se produce si o pra0usire a vascozitatiiKe(ectul senila@rulment !
eritrocitul se muleaza pe capilar si plasma cure.
"resiunea arterialaK tensiunea arteriala ! determinarea se (ace la nivel parietal K
(orta e%ercitata de masa de sane asupra peretilor art su0 in-uenta contractiilor
ritmice ale inimii pentru deplasarea sanelui in ar0orele arterial inc,is. Variatia
presiunii arteriale in sistola VS se inscrie ra+c prin carotidorama. "resiunea
sistolica depinde de volemie$ iar cea diastolica de rezistenta. Tipuri de tensiune
arteriala: "S ma%ima$ presiunea cu care sanele este propulsat in sist art in
sistola 82H-8CHmm=Y ! val normale$ "D minima$ pres cu care sanele continua
sa se deplaseze in ar0orele arterial in diastola ventriculara$ valoare normala X
din "S 48. /n circulatia pulmonara reimul presional este de 8@? din "A. "resiunea
medie !"M ! (ondul presional permanent de propulsie si iriare a sanelui de la
inima la tesut ! G?-8HH. "resiunea pulsului este in-uentata de 2 (actori ! de0it
sistolic si complianta ar0orelui vascular. Amortizarea pulsului spre peri(erie este
determinat de rezistenta la deplasarea sanelui in vase. Variatii ale presiunii
arteriale: varsta 83H-8LHmm=Y creste la 0atrani. 9a copil 8C zileKLHmm=$ 8H
aniK8HHmm=. /n somn: DH@?H mm=((ara vise). "resiunea sistolica poate
aBune la 2HH mm=. <(ort 83H-8DH@3Hmm= (presiunea diastolica e constanta
sau scade usor). <motii stres presiunea arteriala creste. 9a variatii de
temperatura presiunea arteriala scade. /n perioada diestiei presiunea art creste
pe seama volemiei. Yravitatie ! presiunea arteriala creste in artera de su0 cord si
scade in artera de deasupra cordului. 6espirata in inspir creste in e%pir scade.
83
?G.viteza de circulatie$curerea laminara si tur0ulenta
Viteza de circulatie a sanelui este dirept proportionala cu depitul si invers
proportionala cu sectiunea. 9eea lui Eernoulli: cu cat creste viteza de curere cu
atat scade presiunea laterale ce destinde vasul. /ntr-un vas cu placa de aterom
creste viteza de deplasare$ scade presiunea de distensie si permite
permanentizarea o0structiei.viteza scade in capilare si creste in venele cave.
5urerea laminaraKcurerea in straturi. C$L la 0ar0ati$ C$A la (emei si 8$D3 in
plasma. "roprietatile unui lic,id de a si sc,im0a treptat (orma datoritara (recarii
dintre elementele +urate. /mportanta in curerea sanelui: din intermitenta
devine continua si in rezistenta. 6ezistenta la curere a sanelui este de A ori mai
mare ca a apei si este nevoie de o presiune de A ori mai mare de impinere. <%:
anemii: viteza de circulatie a sanelui creste pt ca crete si (recventa cardiaca si
apare ta,icardia ! suprasolicitarea inimii. Sanele cure in ritm constant $ in
planuri paralele. #iecare strat de sane se mentine la distanta eala de peretele
vascular. Viteza in centrul vasului este mai mare decat in straturile marinale:
intre centrul si peretele vasului e%ista multe straturi de molecule care aluneca
intre ele iar moleculale a-ate in contact cu peretele se misca reu datorita
derentei la perete.
5urerea tur0ulenta ! cand viteza de circulatie a sanelui creste$ la nivelul
supra(etei ruoase$ stenoze$cuduri$ depasind un pra critic se trans(orma in
curere tur0ulenta: sanele cure perpendicular pe vas in loc sa cura de-a
lunul luiK varteBuri care reduc viteza sanuina ! su-uriKmani(estare
stetacustica. Tipul de curere poate + apreciat prin numarul lui 6eTnolds cu
valoare normala su0 8HH. /n e(ort +zic viteza sanelui creste. 5and -u%ul de
sane ocoleste un o0stacol in interiorul vasului$ cand isi sc,im0a directia 0rusc
sau ocoleste o supra(ata ruoasa. 5urere transversala a sanelui in interiorul si
de-a lunul vasului se (ormeaza varteBuri ! curenti tur0ionari si produ o rezistenta
mare la curere. /n sementul pro%imal al aortei nr lui reTnolds creste in (aza
rapida de eBectie K tur0ulente (si in sementul pro%imal art pulmonare) in vasele
mici nr lui 6eTnolds scade si nu apar niciodata tur0ulente.
3H.rezistenta vasculara
un numar mai mare de vase sanuine paralele usureaza -u%ul s deoarece
+ecare vas paralel asiura o conductanta suplimentara -u%ului sanelui-
rezistenta totala este mult mai mica decat rezistenta unui sinur vas. 5reste (orta
care propulseaza sanele si scade rezistenta vasculara.
6ezistenta vasculara ! di+cultatea la curere intampinata de orice -uid ast(el
curerea se (ace cu consum de enerie$ iar plata se (ace in unitati de presiune
consumate pe parcurs. 5ea peri(erica se calculeaza$ nu se poate masura. Se
e%prima in dTne sec@cm sau unitati de rezistenta peri(erica (M6").K opozitia la
curere a unui lic,id a-at su0 radient de presiune de 8mm =. 8urp in
,ipertensiune pulmonara.
6ezistenta arteriolara ! la acest nivel rezistenta e ma%ima.
6T"- rezistenta totala peri(ericaK83HH-8LHH dTne
6 aortei si art mariK 8?H dTne
6 arteriolaraK8HHH dTne
6 capilaraK2HH dTne
6 venoasaK8?H dTne. 9a nivel de vas individual rezistenta art e mai mica decat
rezistenta cap. Se produce si o pra0usire a vascozitatiiKe(ectul senila@rulment !
eritrocitul se muleaza pe capilar si plasma cure.
38.presiunea arteriala:de+nitie$parametrii$variatii$metode de determinare
"resiunea arterialaK tensiunea arteriala ! determinarea se (ace la nivel parietal K
(orta e%ercitata de masa de sane asupra peretilor art su0 in-uenta contractiilor
ritmice ale inimii pentru deplasarea sanelui in ar0orele arterial inc,is. Variatia
presiunii arteriale in sistola VS se inscrie ra+c prin carotidorama. "resiunea
8L
sistolica depinde de volemie$ iar cea diastolica de rezistenta. Tipuri de tensiune
arteriala: "S ma%ima$ presiunea cu care sanele este propulsat in sist art in
sistola 82H-8CHmm=Y ! val normale$ "D minima$ pres cu care sanele continua
sa se deplaseze in ar0orele arterial in diastola ventriculara$ valoare normala X
din "S 48. /n circulatia pulmonara reimul presional este de 8@? din "A. "resiunea
medie !"M ! (ondul presional permanent de propulsie si iriare a sanelui de la
inima la tesut ! G?-8HH. "resiunea pulsului este in-uentata de 2 (actori ! de0it
sistolic si complianta ar0orelui vascular. Amortizarea pulsului spre peri(erie este
determinat de rezistenta la deplasarea sanelui in vase. Variatii ale presiunii
arteriale: varsta 83H-8LHmm=Y creste la 0atrani. 9a copil 8C zileKLHmm=$ 8H
aniK8HHmm=. /n somn: DH@?H mm=((ara vise). "resiunea sistolica poate
aBune la 2HH mm=. <(ort 83H-8DH@3Hmm= (presiunea diastolica e constanta
sau scade usor). <motii stres presiunea arteriala creste. 9a variatii de
temperatura presiunea arteriala scade. /n perioada diestiei presiunea art creste
pe seama volemiei. Yravitatie ! presiunea arteriala creste in artera de su0 cord si
scade in artera de deasupra cordului. 6espirata in inspir creste in e%pir scade.
Metode de determinare a "A ! directe Kintravasculara ! introducerea unui cateter
in art si conectarea la un sistem de masurare si indirecte ! palpatorie$
auscultatorie$oscilometrica.
3A.spatiul interstitial
"e distanta scurte dintre capilare si celule di(uziunea prin el permite transportul
rapid al moleculelor mici$ electrolitilor$ su0st nutritive$ reziduurilor celulare$
o%ienului si dio%idului de car0on.
#ormat din (ascicule de colaen si el tisular ce (ormeaza o retea +na in care se
+%eaza ultra+ltratul plasmatic. Di(uziunea prin el ! miscare cinetica !molecula
cu molecula. "e lana lic,idul din el e%ista suvoaie si mici vezicule de lic,id li0er
prezenta 8R in tesutul normal. /n edeme acestea se e%pansioneaza si reprezinta
X din lic,idul interstitial. "resiunea lic,idului interstitial este pozitiva in oranele
capsulara.
8)metoda implantarii de capsule per(orate K- 3mm=. "resiunea neativa
interstitiala contri0uie la learea si consolidarea tesuturilor. /n cavitati naturale
din oranism unde e%ista lic,id li0er valorile sunt neative : spatiul intrapleural:-
Dmm=$ sp epidural:-C si -3 mm=.
Masurarea presiunii lic,idului interstitial li0er cu tampon de 0um0ac -Z -A-Z-
8mm=. "resiunea lic,idului interstitial este cu cativa mm= mai neativa decat
presiunea care inconBoara +ecare tesut
5ompozitia lic,idului interstitial ! proteine A-? @dl$ lipide 8-2@dl (dupa pranz).
5uloare al0uie si lactescenta dupa pranz
3C.Sc,im0ul capilar ! (ortele starlin si di(uziunea
3?. presiunea capilara ! valori metode de determinare
8.canulare directa a capilarelor K metoda micropipetarii: AH-CHmm= la capat
arterial. 8Lmm= la capat nervos
2.metoda izoravimetrica$ det de YaTton K 8Lmm= K presiunea cap medie
(unctionala K H$A. 5and presiunea creste cu peste 2H mm= apare o +ltrare de
3D ori mai mareK edeme ! sistemul lim(atic nu (ace (ata.
33.e(ecte clinice si +zioloice ale sc,im0ului capilar-tesut
<demele ! compromiterea vidului interstitial. 5reste presiunea ,idrostatica !
staza venoasa$ in ,ipoal0uminemie(a( ,epatice scade sinteza si a( renale cresc
eliminarile)$ 0locaB lim(atic$ creste permea0ilitatea capilara ! in-amatii$ arsuri.
Aplicatii +zioloice:
8. 1e(ron ! +ltrare in tot capilarul ! pct izos+mic deplasat la in+nit
2. Alveolele:rea0sor0tie de-a lunul capilarelor$ pct izos+mic deplasat la
zeroKalveole uscate.
8D
5reste permea0ilitatea capilara su0 in-uenta su0st vasodilatatoare !
celulele endoteliale lasa spatii li0ere intre plasma si mem0rana 0azala.
3L.circulatia lim(atica ! structura$ rol$ parametrii
6ol de drenaB$ transport si aparare.
DrenaB ! datorita marii permea0ilitati preia proteine si lipide ce le trece in sane.
Transport- constituentii lic,idului interstitial si su0st nutritive resor0ite in
mucoasa astro intestinala mai ales lipidele
Aparare ! distrue corpii straini.
5apilarele lim(atice lipsesc in S15 si oase. Au diametrul de 8H-?Hum$ marinile
celulelor endoteliale raman -otante$ suprapunandu-se peste o parte din celula
invecinata K valve. 5elulale endoteliale +%ate de tesutul conBunctiv prin
+lamentele de ancorare cu proprietati contractile.
6ata -u%ului lim(atic ! in repausK82H ml@, (2ml@min)K8@8H din +ltrarea la nivel
capilar . 6ata este determinata de 2 (actori:
- "resiunea lic,idului interstitial si radul de activitate a pompei lim(atice.
"resiunea lic,idului interstitial are -u% lim(atic scazut. "ompa lim(atica K
umplerea unui sement dintre 2 valve va determina contractia acestuia$
lic,idul va + pompat in sementul urmator. "ompa este in-uentata de
contractia musculara si de miscarea partilor oranismului$ de pulsatiile
arterelor si de comprimarea tesutului prin o0iecte situate e%tern. "ompa
lim(atica este activa in e(ort. /n repaus are redispozitie la staza si edeme.
3D prezentati rolul de rezervor al venelor
Venele au rol de transport al sanelui neo%ienat de la peri(erie la inima. <ste
depozit si rezervor de sane$ apro%imativ L?Hml sane venos ! 2DHH-A?HHml).
>rane cu rol de rezervor: splinaK8HHml sane$ +catKcateva sute de ml$
venele a0dominale mariKAHHml sane$ ple%uri nervoase su0cutanateKAHH-
D?Hml sane.
5ordul si plamanii ! prin stimulare simpatica isi pot micsora volumul. 5ordul
contri0uie cu ?H-8HH ml si venele pulmonare cu 8HH-AHH ml sane.
6ezervor maBor de saneKteritoriu spla,nic ! activ prin stimulare simpatica si
pasiv prin elasticitatea venelor.
/n e(ort splina este rezervor de sane. 5a si ple%urile venoase su0cutanate $
avand rol si in termorelare.
3G. parametrii circulatiei venoase
D5KDartKDcapilar
6ezistenta vv micaK 8H dTne sec cm?
"resiunea vv peri(erica: la capat vv aS caoK8Hmm=$ restul de " imprimat de
VS. variaza cu pozitia$ in mers nu creste mai mult de 2?mm=$ iar in vene
mariK A-C mm=$ ADK Hmm=.
D5 este corelat cu (unctia de rezervor de sane al venelor si de intoarcerea
venoasa (presarcina si postsarcina). /ntoarcerea venoasa este in-uentata de
suma -u%urilo sanv locale. Vasele se dilata creste intoarcerea venoasa si D5-
Z testul primeste mai mult sane. "resiunea de umplere sistemicaK presiunea
inreistrata in orice parte a circulatiei cand inainteaza orice -u% circulator. 5u
cat presiunea de umplere este mai mare si ste venos este mai plin. #actori
care cresc presiunea de umplere sistemica
- 5reste vol sanuin cu 8?-AHR$ du0leaza p de umplere$ creste contractia
musculara si presiunea pana la 2?mm=. #actori care scad presiunea de
umplere
LH. presiunea venoasa$ tipuri si valori
"V se masoara in AD si nu variaza cu e(ort si pozitie$ daWr variaza cu (azele
ciclului cardiac$ aBune pana la Lmm=.cand nu e%ista intoarcere venoasa$
presiunea venoasa este Lmm=. 5u cat presiunea de umplere este mai
maer si sit venos este mai plin. "resiunea venoasa se masoara in AD (H la
8G
intersectia tricuspidei)$ nu variaza cu e(ort si pozitie$ dar variaza cu (azele
cicluluin cardiac. ABune pana la L mm =$ -A-? mm=. 6ezultatul
ec,ili0rat dintre intoarcerea venoasa si (unctia de pompa. "arametru (olosit
in insu+cienta cardiaca stana ! ,ipertro+a VS$ cresterea presiuniii de
umplere in AD
Masurarea presiunii venoasa
8. Metoda directa ! saneranda ! manometre cu =2>
2. Metoda indirecta ! manometrul recSlin,ausen. are la 0aza principiul
compresiei cu valoare cunoscuta a uneia din venele super+ciale cu
manson de cauciuc. "resiunea peri(erica variaza intre L-8?. /n
insu+cienta cardiaca CH-3H cm =2>.
L8 #actorii intoarcerii venoase
Di(erenta de presiune dintre capetele tu0ului: VS-AD prin mecanismul de
impinere din urma a sanelui. Aspiratia cardiaca ! pompa realizata de VD in
sistola$ 0ene+ciara +ind vena cava in(erioara. Aspiratia toracica ! inspir@e%pir. /n
inspir presiunea intratoracica su0atmos(erica (avorizeaza intoarcerea venoasa.
"resa pozitiva a0dominala ! sanele trece din reiunea mem0relor in(erioare
catre inima$ presiunea venoasa e%traa0dominala depaseste presiunea
a0dominala. Yravitatia ! venele de deasupra cordului sunt cu valoare neativa
dedesu0tul cordului. "ompa musculara ! contractiile izotonice ale musculaturii
sc,eletice (avorizeaza circulatia venoasa ! val presiunii venoase K 2Hmm=. in
ortostatism prelunit "V la e%tremitati aBune la GHmm= in AH secunde.
L2. -u%ul sanuin muscular
6elare prin controlul local al rezistentei vasculare
- De repaus A-Cml@min@8HH
- /n e(ort creste de peste 2? ori K ?H-DHml@min@8HH
- 5apilarele musculare sunt desc,ise in procent de 2H-2?R$ in e(ort +zic
creste -u%ul sanuin datorita mecanismelor locale de relare (relarea
microcirculatorie) si mecanisme nervoase.
Adaptarea circulatiei se (ace prin
- Stimularea S15
- 5resterea presiunii arteriale
- 5restrea de0itului cardiac
Stimulare snc ! semnale de la creier spre centrul vasomotor pt descarcare
simpatica si simultan semnale parasimpatice spre inima$ diminua: inima
determina cresterea (recventei cardiace si e(ort inotrop$ vasoconstrictia
arteriolara peri(erica si vasodilatatia musculara$ vasoconstrictie vv
determina crestrea intoarcerii spre atriul drept.
5restrea presiunii arteriale cu 2H-DHmm= ! vasoconstrictie arteriolara$
cresterea activitatii de pompa$ venoconstrictie importanta
5restrea de0itului cardiac ! prin e(ort +zic se realizeaza predominenta
simparitca cu crestrea de0itului cardiac
LA. (actori determinanti ai de0itului coronarian
5irculatia coronarianaK2?HH cap@mmA. #lu%ul sanuin de repaus K 22?ml@min !
DHml@8HH K C-?R. De0itul cardiac in e(ort creste de C-L ori$ aBunand la 8HHH-
82?Hml@min.
Vasele su0epicardiceKlari$ distensi0ile$ vase de conductanta. Vasele per(orante
si arteriolele ! vase de rezistenta. 5irculatia coronara terminala si colaterala.
#actori care determina de0it coronarian
- "resiunea de per(uzie a coronarelor
- 5ompresia sistolica
- 5onsumul de H2 miocardic
- Su0st vasoactive si tonusul neuroen
2H
- Miocardul e%trae pana la LHR din H2 din sae ! ceea ce duce la cresterea
-u%ului coronarian.
"resiunea de per(uzie K depinde de presiunea sistolica si presiunea
diastolica. /n controlul izovolumetric presiunea ventriculului stan este mai
mare decat presiunea din aorta. De0itul coronarei drepte este eal in
sistola si diasistola ! creste constant cu diasistola datorita presiunii
intraparietale mai mici decat in aorta.
5irculatia s in coronare se (ace datorita duratei diastolei.
5ompresia sistolica ! in stenoza aortica compresiune severa. 6ezistenta pe
care aorta o con(era tre0uie invinsa de o presiune mult mai mare catre
turteste vasle intramurale. 5resterea necesarului de o%ien pentru
contractie ! insu+cienta coronariana
LC de0itul sanuin cere0ral
De0itul sanuin cere0ral mediu K ??-3Hml@8HH@minK L?H-DHHml@min. 5onsumul
de o%ien pentru creierul total este 2HRKCH-?Hml@min
Sursa de enerie ! lucoza (C$Gm@8HH@min)$ in cantitati scad ac ,idro%i0utiric si
acetoacetic. Dintre aminoacizi se sintetizeaza ac lutamic si aspartic. 5ontrolul
umoral creste concentratia de dio%id de car0on si ,idroen. 6elarea
e%tracere0rala intervina cand tensiunea arteriala scade cu ?HR din normal ceea
ce duce la raspunsuri neuro re-e%e compensatorii$ in e(ort ! vasospasm cere0ral
cu rol protector.
L?.#actori inotropi pozitivi
#unctia inotropa (contractilitatea)
Z de+neste proprietatea miocardului de a se contracta ca urmare a stimularii
electrice. contractia optima se (ace la o lunime optima a +0relor de 2$2[
(actori inotropi (4): adrenalina$ \ aonistii adrenerici$ sistemul nervos veetativ
simpatic$ tonicardiacele (diitalice$ 0ipiridine$ metil%antine - teo+lina$
amino+lina)$ 5a
L3.#actori inotropi neativi
(actori inotropi (-): acetilcolina$ \ 0locantele adrenerice$ sistemul nervos
veetativ parasimpatic$ 0locantele canalelor de 5a (D="$ 0enzotiazepine$
(enilalc,ilamine)$ ,iperU$ isc,emia locala (prin cresterea concentratiei de
protoni).
LD.Mecanism mioen de relare a microcirculatiei
"rocesul de autorelare a circula'iei este prezent n maBoritatea oranelor (creier$
inim i rinic,i):
la creier -u%ul sanuin este men'inut constant c,iar dac TA se modi+c (ntre
3H-8DH mm=) )men'ine ec,ili0rul con'inut-con'intor
la rinic,i -u%ul sanuin este men'inut constant c,iar dac se modi+c TA (ntre
8HH-8DH mm=) )
men'ine +ltrarea lomerular constant (#YK82H 8? ml@min).
LG.Mecanisme meta0olice de relare a microcirculatiei
=iperemia activ const n vasodilata'ie cu creterea -u%ului sanuin local n
'esuturile active.
mecanismul de producere:
- intensi+carea activitatii tisulare si a eli0erarii de meta0oliti$ vasodilatatie
locala.
=iperemia reactiv apare la reluarea -u%ului sanuin ntr-o zon n care -u%ul de
s&ne a (ost oprit pentru secunde@minute (e%: aplicarea unui arou pe 0ra').
caracteristici:
- -u%ul sanuin: crete direct propor'ional cu durata ocluziei i revine lent la
valorile ini'iale.
28
cauza o reprezint ac'iunea vasodilatatorie a(actorilor meta0olici locali
=ipo%ia ac'ioneaz direct i prin meta0oli'ii vasodilatatori (adenozina$ acidul
lactic
Adenozina
<ste sintetizat n celulele tisulare$ din AM" n lipsa >2 i di(uzeaz spre vase$
unde ac'ioneaz pe receptori speci+ci-purinerici (localiza'i pe +0rele musculare
netede din peretele vascular
Scderea adenozinei se realizeaz printr-un mecanism de (eed-0acS$ care
readuce vasul la dimensiunile ini'iale
D2 ! e(ectul sistemului nervos simpati asupra relarii -u%ului sanuin
5and -u%ul sanuin scade la nivelul centrului vasomotor din trunc,iul cere0ral
in(erior ! isc,emie cere0rala. 1euronii de la acest nivel raspund direct devenind
e%citati iar presiunea arteriala sistemica creste la ma%imul la care pompa
cardiaca (ace (ata ! into%icarea centrului vasomotor cu dio%id de car0on (4ac
lactic si alti meta0oliti). 5resterea presiunii arteriale ca raspuns la isc,emia
cere0ralaKraspunsul snc la isc,emie.
/mportanta snc isc,emic:
-re-e%ulKactiv la "A su0 3-mm= si e+cient la "A de 8?-2H mm =.
6e-e%ul 5us,inK tip de raspuns al S15 isc,emic ca urmare a cresterii presiunii
arteriale in cutia craniana . cand presiunea 956 creste la valoarea "A$ comprima
arterele cere0rale si opreste -u%ul s cere0ral producand isc,emie K se reinitiaza
raspuns al S15 la isc,emie care determina cresterea presiunii arteriale. 5and
presiunea arteriala e mai mare decat presiunea 956 sanele incepe sa cura prin
arterele cere0rale inlaturand isc,emia.
DA.6eceptori implicati in relarea cardiovasculara
6olul centrilor nervoi superiori n relarea 5V
8. 5entrii din (orma'iunea reticulat ponto-mezence(alic induc e(ect 4@- asupra
centrilor 0ul0o-pontini.
2. =ipotalamusul controleaz centrii 0ul0o-pontini n timpul e(ortului +zic$
emo'iilor$ varia'iilor termice$ al actelor comportamentale (alimenta'ie$ aprare$
activitate se%ual).
=ipotalamusul anterior are neuroni parasimpatici )stimulare ]ona depresoare 5V
) 7#5 4 vasodilata'ie.
=ipotalamusul posterior are neuroni simpatici )stimulare ]ona presoare 5V )
I#5 4 vasoconstric'ie
A. Sistemul lim0ic intervine n controlul centrilor 0ul0o-pontini mpreun cu
,ipotalamusul n strile
emo'ionale. Di(eritele zone pot stimula zona depresoare sau presoare.
C. Talamusul controleaz centrii 0ul0o-pontini in-uen'&nd #5.
?. 5ere0elul stimuleaz activitatea S1VS$ duc&nd la creterea #5$ n condi'iile
adaptrii circula'iei la
modi+crile posturale.
3. 5orte%ul$ n special ariile din Bumtatea anterioar au rol n controlul activit'ii
cardio-vasculare
5entrii 0ul0o pontini cu rol in relarea cardiovasculara:
]ona presoare$ localizat n por'iunea dorso-lateral$ este zona
cardioacceleratoare i vasomotorie:
- controleaz activitatea neuronilor simpatici medulari
i medulosuprarenala.
- stimularea zonei presoare determin un re-e% presor:
I#5 i I#or'ei de contrac'ie cardiace ) I D5.
I tonusului vascular (vasoconstric'ie) ) I rezisten'ei
peri(erice totale (6"T)
22
]ona depresoare - localizat n por'iunea ventro-median$ este zona
cardioin,i0itoare:
cuprinde nucleul dorsal al vaului (1DV) i nucleul am0iuu$ locul de emeren'
a nervilor vai.
Stimularea zonei depresoare determin in,i0area zonei presoare i stimularea
vaal$ duc&nd la un
re-e% depresor
DC.Mecanismul stimularii 0aroreceptorilor
6olul 0aroreceptorilor din sinusul carotidian i crosa
Aortei stimularea lor: dat de distensia pere'ilor arteriali ca urmare a creterii
tensiunii arteriale.
rezultat: re-e% depresor$ cu 7#5 i vasodilata'ie$ duc&nd la 7tensiunii
arteriale:calea a(erent este reprezentat de nervul /^ (loso(arinian) i ^
(va).centrii cardio-vasculari sunt localiza'i n zona 0ul0o-pontin.calea e(erent:
stimularea vaal spre inim iscderea tonusului simpatic pe vase
D3.Su0stante vasoconstrictoare
5atecolaminele secretate de medulosuprarenal:
- adrenalina (A) i noradrenalina (1A) - n mod normal se eli0ereaz n
cantit'i sczute$ cu un e(ect limitat. la stres$ eli0erarea de catecolamine
este masiv )e(ect semni+cativ$ cu modi+carea -u%ului sanuin n
(unc'ie de tipul i de numrul de receptori adrenerici
1oradrenalina ac'ioneaz pe receptorii -adrenerici vasculari
)V5(vasoconstrictie)
Adrenalina ac'ioneaz pe receptorii producand vasodilata'ie n teritoriul
coronarian$ muc,ii
sc,eletici i cere0ral.Am0ele catecolamine activeaz centrii cardiovasculari din
0ul0 i punte
Dopamina -este precursor al 1A.ac'ioneaz pe receptorii -adrenerici vasculari i
produce vasoconstric'ie$ (r modi+carea 6"T.pe receptorii miocardici )e(ect
inotrop i
cronotrop pozitiv
1europeptidul ; (1";) se descarc concomitent cu 1A i are un e(ect
vasoconstrictor puternic i prelunit.
5atecolaminele au o durat de ac'iune sczut pentru c sunt inactivate rapid
enzimatic (de MA> !
monoaminoo%idaze
Vasopresina sau ,ormonul antidiuretic (AD=)produs de ,ipotalamus i depozitat
n ,ipo+za posterioar.are e(ect V5 doar la doze mai mari dec&t cele
o0inuite.rolul su principal este de relare a eliminrii de ap la nivel renal$
pentru men'inerea osmolarit'ii i a volemiei
=ormonii tiroidieni - secreta'i de tiroid -poten'eaz e(ectul catecolaminelor.
(riul I eli0erarea TA$ TC ) V5.
la ,ipertiroidieni$ prin e%aerarea e(ectelor catecolaminelor$ apar e%trasistole$
ta,icardie p&n la
+0rila'ie atrial i crete TA
Mineralocorticoizii: Aldosteronul este secretat de corticosuprarenal nu are e(ect
direct asupra tonusului vascular.
- are e(ect indirect de I a sensi0ilit'ii vaselor la stimuli vasoconstrictori prin
I rea0sor0'iei de 1a4i ap
#actorii endoteliali vasoconstrictori:
- Trom0o%anul A2provine din lipidele mem0ranare. Are rol vasoconstrictor i
areant plac,etar.
2A
- <ndotelina sau (actorul constrictor derivat din endoteliu (<D5#) are rol
vasoconstrictor puternic$ asemntor cu aniotensina // i mult mai
puternic dec&t 1";. <li0erarea <D5# crete la (ri. /ntervine n patoeneza
aterosclerozei$ =TA$ cardiopatiei isc,emice
DL.Su0stante vasodilatatoare
5reterea produc'iei de acid lactic$ =4i 5>2din 'esuturile active (cu meta0olism
crescut i cu p>
2sczut) induce vasodilata'ie local (dar de scurt durat).
5reterea concentra'iei de U4$ a (os(atului anoranic i a osmolarit'ii pot induce
vasodilata'ie local$ dar de scurt durat.
"rostalandinele induc vasodilata'ie local
To'i aceti meta0oli'i au rol n ,iperemia activ dar nici unul nu o poate produce
sinur ci numai n asociere cu al'i (actori.
"rostalandinele - din meta0olizarea acidului ara,idonic din mem0rana celulelor -
inclusiv a celulelor endoteliale. Ac'ioneaz doar local (sunt rapid inactivate n
circula'ie) )VD i in,i0 arearea plac,etar
1ici o su0stan' vasodilatatoare nu s-a sit n cantitate su+cient de mare pentru
a produce sinur creterea -u%ului sanuin n (unc'ie de necesit'ile meta0olice.
Acest proces este rezultatul ac'iunii sinerice a mai multor su0stan'e
vasodilatatoare
:n condi'ii patoloice$ o0struc'ia lumenului vascular (ateroscleroza) scade
capacitatea vasodilatatorie
n condi'iile unui necesar meta0olic crescut (e%. ne(ort)$ c,iar dac crete sinteza
de meta0oli'i locali$ nu se mai poate o0'ine o vasodilata'ie corespunztoare i
consecutiv -u%ul sanuin local scade.
6ezultatul acestei reduceri a -u%ului sanuin este dezec,ili0rul ntre cererea i
o(erta de >2
i su(erin'a tisularla nivelul inimii apare anina pectoral.
la mem0rele in(erioare apare claudica'ia intermitent !durerea n am0e n
timpul mersului
Su0stan'ele 0ioloic active - secretate n 'esut$ ac'ioneaz local$ produc&nd VD
local i Ipermea0ilit'ii vasculare$ cu (ormarea edemului local:
a) =istamina (=/ST)
0) Serotonina
c) Uininele
=istamina (=/ST)
- stocat n ,ipotalamus$ eozino+le$ 0azo+le i mastocite.
- produs prin decar0o%ilarea ,istidinei.
- eli0erare su0 in-uen'a unor stimuli imuni sau neimuni )deranularea
mastocitelor cu eli0erarea de =/ST
-eli0erarea masiv de ,istamin (n alerii) ) oc ,istaminic cu pr0uirea TA.
- n re-e% de a%on: se eli0ereaz i su0stan'e tip =/ST
Serotonina
- I n mastocite$ trom0ocite$ cel. croma+ne astro-intestinale.
- provine din meta0olismul tripto(anului.
- e(ecte: - VD muscular i cutanat (e%tremitatea ce(alic)$
- V5 pe arteriolele i venulele splan,nice
Uininele: 0radiSinina i lisil0radiSinina
- rezult din Sininoen su0 ac'iunea Salicreinei. Activarea preSalicreinei
plasmatice inactive n Salicrein este realizat de (actorului ^// (=aeman).
- sistemul Sininelor este inactivat de dou Sinaze$ dintre care una este c,iar
enzima de conversie
enzima de conversie in,i0 sistemul Sininelor (VD) i stimuleaz sistemul renin-
aniotensin (V5) )
2C
conectarea ntre sistemele vasoconstrictor i vasodilatator local.
- principalele e(ecte ale Sininelor :
- VD arteriolar (puternic dar de scurt durat)$
- creterea permea0ilit'ii capilare$
- (ormarea edemului$
- rol proin-amator$
- relarea -u%ului sanuin n
- piele$
- lande salivare$
- lande sudoripare$
- lande astro-intestinale
"eptidul atrial natriuretic (A1").
- +0rele musculare din atrii au ranule asemntoare cu celulele landelor
endocrine care secret ,ormonii polipeptidici.
- A1" este secretat ca un prepro,ormon$ care se trans(orm ntr-un propeptid
stocat n ranule. Din acest propeptid se eli0ereaz A1" (23 AA) ca (orm
circulant n s&ne.
- A1" se eli0ereaz n urma stimulrii receptorilor atriali de ctre creterea
volemiei$ a 1a4
plasmatic$ n cazul e%cesului de aniotensin // circulant i la creterea #5.
- alimentele cu pu'in 1a4in,i0 eli0erarea de A1"
RESPIRATOR
88.volume si capacitati pulmonare defnitie, valori
Volumele pulmonare statice. Aceste volume cuprind aerul care poate + introdus
sau scos din plm&ni n cursul unei respira'ii. <le sunt (oarte varia0ile c,iar la
normali n (unc'ie
de v&rst$ se% sau a0aritul individului.
Volumul curent (V5) este volumul de aer ce poate + introdus i scos din plm&ni
n cursul
respira'iei de repaus i are o valoare de apro% ?HH ml.
Volumul inspirator de rezerv (V/6) este volumul de aer ce poate + introdus n
plm&ni
la s(ritul unei inspira'ii normale. Are o valoare de A?HH-ADHH ml reprezent&nd
cam 3HR din
capacitatea vital.
Volumul e%pirator de rezerv (V<6) este volumul ma%im ce poate + eliminat la
s(&ritul
unei e%pira'ii normale. Are o valoare de 8HHH-82HH ml sau 2?R din capacitatea
vital.
5apacitatea vital (5V) reprezint suma V5$ V/6 i V<6 deci cam C?HH-CDHH ml.
"oate
+ msurat su0 (orma volumului de aer eliminat printr-o e%pira'ie (or'at ce
urmeaz unei
inspira'ii (or'ate. 6eprezint cam L?R din capacitatea pulmonar total (5"T).
Volumul rezidual (V6) este volumul de aer care rm&ne n plm&ni dup o
e%pira'ie (or'at.
Acest volum care nu poate + eliminat este n Bur de 8?HH ml sau 2?R din 5"T.
5apacitatea rezidual (unc'ional (56#) este suma V<6 i V6 i reprezint
volumul de aer
ce se sete n plm&ni la s(&ritul unei respira'ii normale (22HH-2?HH ml sau
?HR din 5"T).
2?
5apacitatea pulmonar total (5"T) reprezint volumul total de aer con'inut n
plm&ni
+ind eal cu suma 5V i V6 (?-3HHH ml)
Scderea 5V su0 DHR din valoarea NidealO este considerat patoloic dar ea
poate +
compensat prin creterea (recven'ei respiratorii care men'ine ventila'ia ma%im
cu condi'ia
unei 0une permea0ilit'i 0ronice. Scderile net patoloice care nu pot +
compensate presu-
pun o scdere a 5V su0 ?H-3HR.
Alterarea volumelor pulmonare tre0uie corelat cu alte date privind (unc'ia
pulmonar
(de0ite ventilatorii$ complian'a pulmonar$ rezisten'a la -u%$ capacitatea de
di(uziune$
concentra'ia sanuin a azelor respiratorii
6espira'ia normal se numete eupnee$ cea accelerat polipnee$ iar cea
ncetinit0radip-
nee. 5reterea amplitudinii respira'iei se numete ,iperpnee iar oprirea
respira'iei apnee
89.capacitatea reziduala unctionala
5resterea ei insoteste de cele mai multe ori pe cea a volumului rezidual$
mecanismul de producere +ind acelasi.scaderea capacitatii reziduale (unctionale
si a volumului rezidual pot + intalnite in (ri0roze pulmonare di(uze si in 0olile
cardiace cu staza pulmonara.
capacitatea reziduala (unctionala (5.6.#.)$ reprezinta cantitatea de aer care
ramane in plaman in cursul respiratiei normale$ constand din V.<.6.(volumul
e%pirator de rezerva8?HHml) 4 V.6.ea este cam ?HR din capacitatea pulmonara
totala.
9!."E#S
Volumul e%pirator ma%im pe secund (V<MS) reprezint volumul ma%im e%pirat n
prima
secund a unei e%pira'ii (or'ate ce urmeaz unei inspira'ii (or'ate. Se determin
spirora+c i
poate + corelat cu cur0a -u%-volum (#/Y 2.8C).
<%primarea V<MS se poate (ace n valoare a0solut (ml)$ aceasta tre0uind s +e
peste DHR
din valoarea ideal sau se poate raporta la 5V su0 (orma indicelui de
permea0ilitate 0ronic
V<MS%8HH@5V care tre0uie s +e mai mare de LH.
V<MS olindete mai ales rezisten'a la -u% n cile aeriene peri(erice de mic
cali0ru care
poate + modi+cat prin procese in-amatorii sau alerice$ dar i scderea (or'ei
contractile a
muc,ilor respiratori. Scderea V<MS se nso'ete (recvent de tul0urri de
distri0u'ie a aerului
ventilat i creterea V6 i a 56#.
Volumul inspirator ma%im pe
secund(V/MS)estevolumuldeaerinspiratnprimasecund
a unei inspira'ii ma%ime rapide care urmeaz unei e%pira'ii ma%ime. Valoarea sa
depete cu
pu'in valoarea V<MS raportul V<MS@V/MS +ind de H.D. V/MS scade mult n
stenozele +%e ale
cilor aeriene produse de o0icei prin leziuni cicatriciale
9$.ortele de recul elastic toraco pulmonare
23
Cutia toracic foia parietal a pleurei
Plmnii foia visceral a pleurei
Plmnii foia visceral a pleurei
Volumul de repaos elastic al cutiei toracice
Volumul de repaos elastic al plmnilor
Consecine formarea de presiune subatmosferic
ntre foiele pleurale vid pleural
Inegalitatea vidului pleural
Pe plmn vertical
clinostatism
9%.vidul pleural ormare, valori
nu este vid real ci presiune subatmosferica
Geneza vidului pleural forte de recul elastic ale cutiei toracice si ale plamanilor
Inegalitatea vidului pe plamanul vertical consecinta inegalitatii vidului ventilatie regionala neuniforma
9&.ine'alitatea re'ionala a ventilatiei
9(.ine'alitatea locala a ventilatiei
9).complianta pulmonara statica
Valoarea normal a complian'ei este de H.2 9@cm ap.
5omplian'a static se determin la volume +%e n a0sen'a -u%ului de aer$ n timp
ce
complian'a dinamic se determin n cursul unui ciclu respirator o0inuit.
6aportarea complian'ei la capacitatea vital (ea scade odat cu volumul
plm&nilor)
reprezint complian'a speci+c.
5omplian'apulmonarcretenem+zemulpulmonarncarerecululelasticscadedatorit
distruerii pere'ilor alveolari. <a scade n a(ec'iunile care duc la +0rozarea
'esutului pulmo-
nar.
5omplian'a toraco-pulmonar este de doar Bumtate (H.8 9@cm ap) din valoarea
celei
pulmonare din cauza riidit'ii mai mari a cutiei toracice
5omplian'a pulmonar e%prim elasticitatea acestuia i poate + apreciat prin
volumul de aer care
poate + introdus n plm&ni pentru o di(eren' de presiune de 8 cm ap. /n
condi'ii normale are o valoare
de H.2 9@cm ap.
5omplian'a toraco-pulmonar are o valoare mai mic de numai H.8 9@cm ap din
cauza unei
elasticit'i mai reduse a toracelui comparativ cu plm&nul.
9*.+isteresis
Modul n care se poate urmri rela'ia volum-presiune (cur0a ,isteresis) pe
plm&nul izolat. Se
msoar varia'iile de volum ale plm&nului ca urmare a modi+crii presiunii n
vasul n care se a- acesta.
/n cazul n care plm&nul este umplut cu o solu'ie salin (deci (r inter(a'a aer-
lic,id) sunt operante numai
(or'ele elastice ale 'esutului (r e(ectul tensiunii super+ciale$ presiunea necesar
distensiei plm&nului +ind
cam de A ori mai mic. Dac ndeprtm sur(actantul volumul pulmonar va + mai
mic la aceeai presiune
9,.complianta pulmonara dinamica
2L
98.tensiunea superfciala alveolara
99.suractantul
Moleculele sur(actantului au un capt ,idro+l spre (aza lic,id i unul ,idro(o0
spre (aza
azoas. Atunci c&nd alveolele se destind n inspira'ie moleculele se
ndeprteaz i ten-
siunea super+cial crete mpiedic&nd supradistensia. #enomenele au loc invers
n e%pira'ie
c&nd alomerarea moleculelor de sur(actant mpiedic cola0area alveolelor
datorit scderii
tensiunii super+ciale.
Sur(actantul se (ormeaz n pneumocitele ranulare celule alveolare specializate
din care
se eli0ereaz su0 in(luen'a distensiei alveolare i a stimulrii F adrenerice.
Sinteza sa este (avorizat de lucocorticoizi$ estroeni$ A5T=$ ,ormonii tiroidieni$
prostalan-
dine$ cAM".
Sur(actantul are mai multe roluri:
-mpiedic supradistensia sau cola0area alveolar i reduce e(ortul respirator
-aBut la recrutarea de noi alveole i la ventila'ia colateral mrind porii
-este par'ial reponsa0il de ,istereza 0uclei volum-presiune
-previne e%travazarea lic,idian n alveole
-aBut la cur'irea alveolelor.
Scderea cantit'ii de sur(actant duce la rave tul0urri respiratorii ce
caracterizeaz
sindromul de detres respiratorie$ mai (recvent la prematuri. 5reterea e(ortului
respirator$
reducerea ventila'iei alveolare$ cu ,ipo%emie i acumularea de 5>2 se nso'esc
de tul0urri
ciculatorii care pot provocamoartea.
Administrareadesur(actantsaupunerea n repaus a plm&nilor p&n la re(acerea
sur(actan-
tului prin o%ie-narea e%tracorporeal de mem0ran (<5M>) poate aButa n ast(el
de cazuri
Sur(actantul pulmonar endoen :Sur(actantul pulmonar endoen determina
tensiunea super+ciala pe supra(ata alveolelor in timpul respiratiei si sta0ilizeaza
alveola impotriva colapsului in timpul presiunii transpulmonare de repaus.
De+cienta sur(actantului pulmonar cauzeaza Sindromul de Detresa 6espiratorie
(SD6) la copii prematuri
$!!.spatiul mort
5ompozitia aerului alveolar tre0uie sa ai0a insa o valoare aproape constanta$
aceasta realizandu-se prin inspiratie$ care (ace sa patrunda aer atmos(eric 0oat
in >2$ aer care se distri0uie si se amesteca cu aerul alveolar. 1u tot aerul
inspirator aBune la alveole$ o parte (circa AHR) ramanand in caile aeriene
superioare. Acesta este spatiul mort anatomic.
cuprinde volumul de aer a-at n cile aeriene i plm&ni care nu particip la
sc,im0urile
respiratorii. <l reprezint cam 8@A din volumul inspirator
cuprinde totalitatea aerului care dei este inspirat nu particip la sc,im0urile
respiratorii. De cele mai multe ori su0 aceast denumire este considerat spa'iul
mort anatomic
reprezentat de aerul din cile aeriene (8?H ml) cu pere'i care nu permit
sc,im0urile de aze.
2D
/n aceste condi'ii din cantitatea de aer inspirat numai 2@A aBune n alveole
constituind
(rac'iunea alveolar. <+cien'a ventilatorie se e%prim prin raportul dintre
ventila'ia alveolar
i cea lo0al i este apreciat cam la LHR.
<%ist ns i un spa'iu mort +zioloic ce cuprinde volumul de aer care dei
ptrunde
n alveole nu particip la un sc,im0 azos e+cient din cauza raportului
ventila'ie@per(uzie
neadecvatprinalterareaoricreiadincomponenteleacestuia
$!$.etapele ventilatiei
"entilatia proces ritmic automat controlat de centrii din tr cere-ral
care pe timp scurt devine proces voluntar si care suera si re'la. umoral
su- in/uenta presiunii arteriale de !%, co% si p+.
Sindromul -lestemul ondinei pierderea controlului voluntar al
respiratiei leziuni la nivelul centrilor nerv primari respiratorii.
0entrii se a/a in -ul- si punte . in -ul- senzori sist lim-ic, control
cortical si in punte cei eectori mm respiratori
$!%.rezistenta la /u1 distri-utie,valori
6ezisten'a la -u%. Trecerea aerului prin cile aeriene nt&mpin o rezisten' de
(recare
datorit interac'iunii moleculelor de az i datorit (recrii aerului de pere'ii
conductelor.
6ezisten'a la -u% reprezint cea mai mare parte a rezisten'ei v&scoase i depinde
n condi'ii de
repaus de volumul pulmonar$ dispozi'ia cilor aeriene$ (azele respira'iei$ reimul
de curere al
aerului (laminar sau tur0ulent) i cali0rul 0ronic.
5ali0rul cilor aeriene (mai ales al celor lipsite de cartilaii) poate + in-uen'at
c,iar la in-
dividul normal de (azele respira'iei. /n timpul inspira'iei presiunea neativ din
spa'iul pleural
duce la distensia cilor aeriene. /n timpul e%pira'iei presiunea transmural
depinde de modul
n care are loc aceasta. /ntr-o e%pira'ie normal ce are loc pasiv cile aeriene
rm&n destinse$
dar n timpul unei e%pira'ii (or'ate care crete presiunea interpleural p&n la
valori pozitive
cali0rul acestora se micoreaz din cauza (enomenului de compresie dinamic
(#/Y 2.D).
5ompresia dinamic limiteaz -u%ul e%pirator mai ales n cazul e%isten'ei
a(ec'iunilor
pulmonare o0structive care se nso'esc i de leziuni ale parenc,imului pulmonar.
Valoarea rezisten'ei totale la -u% este de 8.3 cm ap@9@s din care H.L cm ap@9@s
pentru
cile aeriene superioare i H.G cm ap@9@s pentru cile aeriene in(erioare.
- ci respiratorii superioare ! DHR
ci respiratorii superioare DHR
! curere tur0ulent
P - ci respiratorii in(erioare ! 2HR
! curere laminar
Controlul neuro-umoral al bronhomotricitii
Factori fizici care influeneaz rezistena la flux
presiune transmural
2G
presiune transmural
Compresia dinamic a cilor aeriene punctul de
presiune egal
Respiraie de repaos
Capacitate vital forat
Constanta de timp a plmnilor - = C x R
$!&.controlul -ron+omotricitatii nervos si umoral al -ran+omotricitatii
$!(.punctul de presiune e'ala
$!).compresia dinamica a cailor aeriene
$!*.valori ale presiunii in circulatia pulmonara
/n inspira'ie muc,ii respiratori nvin rezisten'ele pulmonare i mresc volumul
toraco-
pulmonar.<%pira'iaderepausarelocpasivprinrevenireatoraceluiladimensiunilededin
ainte
de inspira'ie ns pentru o e%pira'ie (or'at este nevoie de interven'ia muc,ilor
e%piratori.
#lu%ul de aer prin cile aeriene este determinat de Bocul presiunilor e%ercitate n
di(eritele
teritorii toraco-pulmonaresu0in-uen'amicrilorrespiratorii.
"resiunea intrapulmonareste presiunea e%istent n cile aeriene i alveolele
pulmonare i
esteealnrepauscupresiuneaatmos(eric./ninspira'ieaceastpresiunescadela!
Amm=
iar n e%pira'ie atine 4A mm=. Aceste valori se modi+c mult n respira'ia
(or'at sau c&nd
e(ortul respirator se (ace cu lota nc,is
"resiunea interpleural sau intratoracic se poate msura n spa'iul dintre (oi'ele
pleurale i are o valoare su0atmos(eric de p&n la !? mm= n condi'ii de
respira'ie normal
"resiunea interpleural nu tre0uie con(undat cu presiunea lic,idului interpleural
care este n Bur de !8H sau !8? mm= datorit tendin'ei capilarelor pleurale de a
a0sor0i lic,idul din spa'iul interpleural datorit presiunii ,idrostatice reduse din
capilare (L mm=).
Di(eren'a dintre presiunea intrapulmonar i cea interpleural constituie
radientul de presiune
transpulmonar care variaz continuu n cursul ciclului respirator (avoriz&nd -u%ul
aerului i av&nd in-uen' asupra ntoarcerii venoasepresiunile intravasculare
pulmonare. "resiunea a0solut este dat de sistola ventriculului drept n
condi'iile rezisten'ei sczute a vaselor pulmonare.
/n artera pulmonar presiunea sistolic este de 2H-2? mm= iar cea diastolic de
3-82
mm= cu o presiune medie de 8? mm=. /n capilarele pulmonare presiunea este
de doar 3-G
mm=$ cu mult mai mic dec&t cea din capilarele circula'iei sistemice. Dac un
cateter este
mpinsprinarterapulmonarp&nlaniveluluneiramurimiciperi(erice$presiuneanrei
strat
este (olosit pentru evaluarea presiunii capilare (capillarT _ede pressure).
9a nivelul venelor pulmonare presiunea scade treptat pentru a aBune n atriul
st&n la
A-C mm=
$!,.enomenele de distensie si recrutare in circulatia pulmonara
AH
0irculatia unctionala prezinta doua proprietati2 prezinta 2 proprietati:
distensie (dilatarea vaselor prin cresterea razei de sectiune ca urmare a cresterii
ntoarcerii venoase sau ca urmare a cresterii presiunii n AS) si recrutare (creste numarul
de capilare functionale ca urmare a cresterii presiunii sau debitului sangvin, ex. n efortul
fizic). vasele extraparechimatoase se afla sub influenta variatiilor presionale pleurale: n
inspir, cnd presiunea intrapleurala scade, presiunea transmurala (ntre artera pulmonara
si pleura) actioneaza ca presiune de distensie si creste fluxul sangvin pulmonar
$!8.caracteristici 'enerale in circulatia pulmonara
5irculatia (unctionala (mica circulatie$ circulatia pulmonara)
Z se (ace n reim de Boasa presiune$ cu o viteza (oarte mare si pe o supra(ata
capilara imensa. (cresterea 0rusca a presiunii n artera pulmonara poate duce la
e%travazare n interstitiu si apoi n alveole$ cu aparitia edemului pulmonar acut)
Z reprezinta ntre de0itul VD
Z rol: de +ltru (se retin si se lizeaza trom0usii veniti din circulatia venoasa
sistemica$ se inactiveaza noradrenalina)$ endrocrin (sediul sintezei enzimei de
conversie a aniotensinei /)
Z vasele micii circulatii sunt (oarte compliante$ reception&nd acelasi volum
sanvin (de0itul VD K de0itul VS) ca si circulatia arteriala sistemica
Z reprezinta un sediu al s&nelui de depozit (H$?-2 l)
Z distri0utia rezistentei este apro%imativ ?HR la nivelul capilarelor$ 8H-8?R n
vene si A?-CHR n artere.
Z prezinta 2 proprietati: distensie (dilatarea vaselor prin cresterea razei de
sectiune ca urmare a cresterii ntoarcerii venoase sau ca urmare a cresterii
presiunii n AS) si recrutare (creste numarul de capilare (unctionale ca urmare a
cresterii presiunii sau de0itului sanvin$ e%. n e(ortul +zic).
Z presiunea de per(uzie a lo0ilor pulmonari variaza n raport cu localizarea (ata
de cord:
o presiunea medie n artera pulmonara este de 8? cm col =2>
o 0aza plam&nilor (situata la 8? cm su0 nivelul cordului) va + iriata cu
presiunea dezvoltata de VD 4 presiunea data de naltimea unei coloane
,idrostatice de 8? cm K 8? 4 8? K AH cm col =2>
o ape%ul plam&nilor (situat la D cm deasupra nivelului cordului) va + iriat cu
8?-D K L cm col =2>.
Z vasele e%traparec,imatoase se a-a su0 in-uenta variatiilor presionale pleurale:
n inspir$ c&nd presiunea intrapleurala scade$ presiunea transmurala (ntre artera
pulmonara si pleura) actioneaza ca presiune de distensie si creste -u%ul sanvin
pulmonar. :n e%pir$ -u%ul sanvin este limitat de presiunea pleurala pozitiva.
Z vasele intraparenc,imatoase K capilare pulmonare$ se a-a si ele su0 in-uenta
(azelor respiratiei (n inspir presiunea alveolara pozitiva devine presiune de
compresie asupra capilarelor pulmonare si limiteaza -u%ul sanvin capilar$ n
e%pir -u%ul este eli0erat)
Z ]onele `.E. aest pulmonare descriu relatia dintre presiunea alveolara$
presiunea n capatul arterial si n capatul venular al capilarului pulmonar.
o zona aest (8)$ presiunea alveolara Z presiunea n capatul arterial: -u%ul
este nul
o zona aest (2)$ presiunea n capatul arterial Z presiunea alveolara Z
presiunea n capatul venular: -u%ul este intermitent n raport cu (azele respiratiei
si (azele ciclului cardiac - n inspir sau diastola presiunea la capatul arterial este
prea mica si -u%ul este limitat$ n e%pir sau sistola presiunea la capatul arterial
depaseste presiunea alveolara si -u%ul este eli0erat.
o zona aest (A)$ presiunea n capatul arterial Z presiunea n capatul venular
Z presiunea alveolara: -u% continuu.
o ]ona aest (2) se reaseste la ape%ul plam&nilor$ n rest avem zona aest
(A). ]one aest (A) nu se reasesc n mod normal.
A8
Z 6elarea circulatiei pulmonare:
o pasiva$ prin (enomene de distensie si recrutare capilara
o activa
Z nervos: prin interventia S1VS sau
Z umoral: su0stante vasoconstrictoare (T^A2$ AY //$ endotelina$ 9T$ "Y<$
serotonina$ ,istamina)$ vasodilatatoare ("5$ 1>$ Ac,)
Z ,ipo%ia este stimulul principal care determina vasoconstrictie (n
a(ectiunile pulmonare cronice care determina insu+cienta respiratorie ,ipo%ica
apare ,ipertensiune pulmonara prin vasoconstrictie persistenta) - acest re-e%
arterioloconstrictor este important deoarece n cazul unei o0structii 0ronsice cu
,ipoventilatie$ raport V@V scazut si ,ipo%ie$ vasoconstrictia locala limiteaza
contaminarea cu s&ne insu+cient o%ienat.
Z sc,im0urile capilar - intestitiu sunt pro+ltrare con(orm presiunilor Starlin:
presiune ,idrostatica capilara K L mm= (pro+ltranta)$ presiune ,idrostatica
interstitiala K - D mm= (pro+ltranta)$ presiune coloid osmotica interstitiala K 8C
mm= (pro+ltranta)$ presiune coloid osmotica capilara K 2D mm= (anti+ltranta).
presiunea neta K 2G mm= pro+ltrare - 2D mm= anti+ltrare K 48 mm=
pro+ltrare
$!9.contaminarea venoasa fziolo'ica
5irculatia n plam&n este du0la: una (unctionala - data de mica circulatie (VD -
artera pulmonara - capilare - venele pulmonare - AS) si una nutritiva - data de
arterele 0ronsice$ apartin&nd marii circulatii (circulatiei sistemice). :ntre aceste 2
circulatii e%ista anastomoze (unctionale ntre venele 0ronsice si capatul venular al
capilarelor pulmonare. datorita radientului presional se realizeaza un sunt
dreapta-st&na$ av&nd ca rezultat contaminarea venoasa +zioloica a s&nelui
arterializat. :n cazul os0tructiei@a0sentei circulatiei pulmonare (atrezie de artera
pulmonara sau stenoza pulmonara)$ circulatia 0ronsica este capa0ila prin
anioeneza sa sta0ileasca comunicatii cu capatul arterial al capilarelor
pulmonare ast(el nc&t sa +e pastrat sc,im0ul azos sanvin (circulatia 0ronsica
poate creste p&na la 2HR din de0itul cardiac
$$!.distri-utia /u1ului de san'e in circulatia pulmonara
$$$.vasele pulmonare e1traparen+imatoase
vasele extraparechimatoase se afla sub influenta variatiilor presionale pleurale: n inspir,
cnd presiunea intrapleurala scade, presiunea transmurala (ntre artera pulmonara si
pleura) actioneaza ca presiune de distensie si creste fluxul sangvin pulmonar. !n expir,
fluxul sangvin este limitat de presiunea pleurala pozitiva
$$%.circulatia pulmonara intraparen+imatoasa
5ircula'ia pulmonar sau mica circula'ie se des(oar ntr-un reim de presiune
sczut dar cu un
de0it eal cu de0itul cardiac (? 9@min).
"resiunea arterial medie din artera pulmonar este de 8? mm= iar cea din
capilarele pulmonare
este de L mm=.
6ezisten'a vascular pulmonar este mai mic (a' de cea din circula'ia
sistemic +ind n Bur de 8HH
dTne@s.cm
#lu%ul sanuin pulmonar este ceva mai mare pentru plm&nul drept (?? R)
comparatriv cu cel st&n.
Distri0u'ia sa nu este eal -u%ul +ind mai mic la v&r(uri i mai mare la 0aze.
Volumul sanuin pulmonarreprezint cam 2CH ml@m2 sau G R din volumul
sanuin total.
6aportul dintre ventila'ie i per(uzie (V@") are o valoare medie de H.D +ind mai
mare la v&r(uri (A.A) i mai mic la 0aze (H.3A) din cauz c ventila'ia crete doar
de A ori de la v&r(uri spre 0aze comparativ cu per(uzia care crete de 8H
A2
ori.6elarea circula'iei pulmonare este dependent de ac'iunile neuro-umorale$
dar i de (actori pasivi.
Vasoconstric'ia pulmonar este produs de e%citarea simpatic ca i de
numeroase su0stan'e (catecolamine$ aniotensina //$ ,istamina$ ,ipo%ie i
,ipercapnie alveolar). Vasodilata'ia pulmonar este produs de parasimpatic i
su0 ac'iunea acetilcolinei$ 0radiSininei$ prostaciclinei sau a 1>
5ircula'ia pulmonar se des(oar n condi'ii unice ntr-un oran care con'ine
aer i a crui (orm se sc,im0 mereu su0 ac'iunea Bocului radientului de
presiune transpulmonar care in-uen'eaz condi'iile ,emodinamice.
5ircula'ia pulmonar constituie mica circula'ie n care presiunile sunt mai sczute
dec&t
cele din circula'ia sistemic dei de0itul este eal cu de0itul cardiac.
5ircula'ia pulmonar ncepe n ventriculul drept continu&nd cu artera pulmonar
care se
divide n 2 ramuri pentru cei 2 plm&ni. "ere'ii arteriali sunt mai su0'iri i au un
strat muscular
mai sla0 reprezentat. 5ele 2 ramuri ale arterei pulmonare se rami+c treptat
urm&nd traseul
cilor aeriene p&n la nivelul precapilarelor care aBunse la nivelul 0ron,iolelor
terminale i al
canalelor alveolare se rami+c ntr-o re'ea de capilare (oarte 0oat
Timpul de circula'ie n circula'ia pulmonar este n repaus de C-? s din care
apro%H.Lsn
teritoriul capilar. /n cursul e(ortului acest timp poate scdea p&n la H.A s.
$$&.re'larea pasiva in circulatia pulmonara
$$(.re'larea nervoasa si umorala a circulatiei pulmonare
$$).eectele +ipo1iei asupra circulatiei pulmonare
$$*.circulatia -ronsica
5ircula'ia 0ronic apar'ine marii circula'ii i se des(oar ntr-un reim de
presiune
,idrostatic crescut$ dar cu un de0it redus (8-2R din de0itul cardiac
$$,.raportul ventilatie3peruzie
/nterrela'ia dintre ventila'ia i per(uzia pulmonar. Dac se consider c ntr-
un minut se ventileaz la nivel alveolar cam C 9 de aer (restul rm&n&nd n
spa'iul
mort) iar de0itul cardiac este de ? 9@min atunci raportul dintre ventila'ie i
per(uzie
(V@") are o valoare medie de H.D n condi'ii ideale n care toate alveolele sunt eal
ventilate i per(uzate.
/n realitate e%ist mari varia'ii ale distri0u'iei aerului ventilat i ale per(uziei ceea
ce (ace
ca raportul V@" s varieze ntre A.A la v&r(uri i H.3A la 0aze. Acest lucru
enereaz un radient
vertical al presiunilor par'iale ale >2i 5>2n aerul alveolar
5u toate c p>2n alveole scade cu CH mm= de la v&r( la 0az$ satura'ia
s&nelui ce
prsete plm&nii nu scade cu mai mult de CR datorit (ormei cur0ei de
disociere a >2=0.
9a individul normal inealitatea raportului V@" are in-uen' minim asupra
sc,im0urilor
respiratorii prin interven'ia a 2 mecanisme compensatorii:
-reducerea per(uziei unui teritoriu 0ine ventilat duce la scderea 5>2din alveolele
i cile
AA
aeriene adiacente ceea ce produce constric'ia musculaturii netede dela acest
nivel diminu&nd
ventila'ia
-,ipoventila'ia unui teritoriu 0ine per(uzat scade >2i ca urmare se produce
vasoconstric'ie care scade per(uzia.
$$8.zonele 4est pulmonare
5onele 4est pulmonare descriu relatia dintre presiunea alveolara A,
presiunea din capatul arterei capilare a si presiunea din capatul venos
cap v.
In zona 4est $, presiunea alveolara comprima circulatia si /u1ul este !.
Este zona de electie pentru instalarea T60.
In zona 4est % /u1ul este intermitent, depinde de azele ciclului cardiac
si respiratie.
5ona 4est & vasul este complet destin' si /u1ul este continuu.
In zona 4est ( presiunea din capatul venos este mai mare decat
presiunea alveolara. 5ona 4est ( apare cand presiunea in atriul stan'
creste2 stenoza mitrala, insufcienta ventriculara stan'a -rusc instalata
ceea ce duce la inarct. Principalul mecanism de instalare a edemului
pulmonar.
$$9.coefcient de solu-ilitate,coefcient de diuziune
$%!.compozitia aerului alveolar
5ompozitia aerului alveolar tre0uie sa ai0a insa o valoare aproape constanta$
aceasta realizandu-se prin inspiratie$ care (ace sa patrunda aer atmos(eric 0oat
in >2$ aer care se distri0uie si se amesteca cu aerul alveolar. Acesta este spatiul
mort anatomic. "e de alta parte$ nu toate alveolele sunt ventilate uni(orm$
aproape 2HR +ind ,ipoventilate (portiunile centrale si paraverte0rale) - spatiul
mort +zioloic. Spatiul mort incalzeste si satureaza cu vapori de apa aerul
atmos(eric si asiura o compozitie constanta aerului alveolar.
in situatii patoloice$ cum sunt reducerea cali0rului 0ronsic (astm$ 0ronsita$
compresiuni etc.) sau cola0areaparenc,imului (atelectazie$ pleurezii etc)$
distri0utia aerului in plamani este neuni(orma$ ea determina cresterea zonelor de
alveole ,iperventilate
$%$.actorii care in/uenteaza diuziunea alveolo7capilara
Di(uziunea alveolo-capilar reprezint trecerea azelor din aer n s&ne i invers
prin traversarea
mem0ranei alveolo-capilare.
Di(uziunea depinde n primul r&nd de di(eren'a de presiune par'ial a azelor$ ca
i de propriet'ile
lor +zico-c,imice sau rosimea i supra(a'a mem0ranei alveolo-capilare.
"entru >2presiunea n aerul alveolar este de 8HH mm=$ iar n s&nele venos CH
mm=.
"entru 5>2presiunea n alveole este de CH mm= iar n s&nele venos C3 mm=.
5apacitatea de di(uziune a plm&nilor const n a0ilitatea unui az de a trece prin
mem0rana
alveolo-capilar n unitatea de timp pentru o anumit di(eren' de presiune
par'ial.
5apacitatea de di(uziune pentru > 2este de 8?-2H ml@min@mm= n repaus i de
CH-DH ml n cursul e(ortului
9a nivelul mem0ranei alveolo-capilare are loc sc,im0ul de aze prin di(uziune$
(enomen
care depinde de mai mul'i (actori:
-di(eren'a de presiune par'ial a azelor
-propriet'ile lor +zico-c,imice
-rosimea i supra(a'a mem0ranei de sc,im0
AC
Di(uziunea azelor se (ace n virtutea radientului de presiune par'ial a
o%ienului i a
5>2 de o parte i de alta a mem0ranei alveolo-capilare. S&nele sosit prin
artera pulmonar este
ncrcat cu 5>2 av&nd o presiune par'ial de CL mm =. :n aerul alveolar$ 5>2
are o presiune
par'ial de numai CH mm =. 5on(orm leilor +zice$ 5>2 va di(uza de la
presiunea mai mare din
capilare la presiunea mai mic din aerul alveolar.
:n aerul alveolar >2 se sete su0 o presiune par'ial de 8HH mm = iar n
s&nele capilar
are CH mm = i va di(uza deci din aerul alveolar n s&nele capilar.
Di(uziunea azelor la nivel
pulmonar este (avorizat i de suprafaa mare de schimb (cca. DH m2) a celor
apro%imativ AHH
milioane de alveole pulmonare. Tot un (actor (avorizant l reprezint i
distana de difuziune (oarte
mic$ mem0rana alveolo-capilar av&nd o rosime de H$?-8b. 5oe+cientul de
di(uziune a 5>2 este
de AH de ori mai mare ca cel al >2$ ceea ce e%plic vitezele de di(uziune
practic eale pentru cele
dou aze$ dei radientele de presiune sunt di(erite (3H mm = pentru >2 i
numai L mm =
pentru 5>2).
Di(eren'a de presiune par'ial. #a' de aerul atmos(eric care con'ine o%ien$
0io%id de
car0on i azot ntr-o anumit propor'ie$ aerul alveolar i s&nele care vine sau
prsete
capilarele con'in mai pu'in o%ien i mai mult 0io%id de car0on$ n timp ce azotul
nu su(er
modi+cri importante (Ta0el /).
Aerulalveolarareocompozi'iedi(eritdeceaaaeruluiatmos(ericdatoritsc,im0urilord
e
aze. <a nu rm&ne +% datorit varia'iilor respiratorii$ a e%isten'ei spa'iului mort
i a inealit'ii
ventila'iei alveolare. Moleculele de az traverseaz mem0rana alveolo-capilar
prin di(uziune
simpl de la locurile unde presiunea lor par'ial este mai mare spre locurile unde
aceasta
Di(uziunea este (avorizat de circula'ia intens$ reac'iile c,imice rapide ale
azelor n
s&ne (avorizate de unele enzime speci+ce i aBustarea continu a ventila'iei i
per(uziei la
necesit'ile meta0olice
$%%.actorii care dierentiaza diuziunea 'azelor
"ropriet'ile +zico-c,imice ale
azelor.Moleculeleazelorsea-ncontinumicareceea
ceasiurdezvoltareauneianumitepresiunin(unc'iedenumruldemolecule$volumul
pecare
l ocup i temperatur (r ca presiunea par'ial a unui az s a(ecteze
presiunea altuia.
/n-uen'a temperaturii asupra volumului azelor este important pentru
msurarea
A?
volumelor pulmonare 'in&nd cont de (aptul c temperatura corporal de
ALrade5 este de o0icei
superioartemperaturiiatmos(ericereduc&ndvolumulazelorcareprsescplm&nii.
/nplus
aerul e%pirat este saturat cu vapori de ap la o presiune par'ial de CL mm=.
Volumele de aer msurate tre0uie deci corectate n (unc'ie de temperatur$
presiunea
atmos(eric i presiunea vaporilor de ap
5antitatea de az din plasm sau alte lic,ide ale corpului depinde de presiunea
azului
i solu0ilitatea acestuia.
Solu0ilitatea azelor este mai sczut n plasm din cauza prezen'ei su0stan'elor
solide
dizolvate i a eritrocitelor.
$%&.unitatea respiratorie
$%(.mem-rana de diuziune
Yrosimea i supra(a'a mem0ranei alveolo-capilare. Dei rosimea medie a
mem0ranei
alveolo-capilare este n Bur de doar 8 [m$spa'iul pe care azele tre0uie s-l
str0at din alveol
p&n n eritrocite sau invers este mult mai comple% aici intervenind i o serie de
alte procese
c,imice.
6educerea supra(e'ei de sc,im0 sau alterarea mem0ranei alveolo-capilare n
cursul unor
a(ec'iuni pulmonare nreuneaz ntr-un mod semni+cativ di(uziunea
6educerea supra(e'ei de sc,im0 sau alterarea mem0ranei alveolo-capilare n
cursul unor
a(ec'iuni pulmonare nreuneaz ntr-un mod semni+cativ di(uziunea
$%).ormula 8ic9 aplicata la diuziunea 'azelor
$%*.capacitatea de diuziune a plamanilor
5apacitatea de di(uziune a plm&nilorconst n a0ilitatea unui az de a trece
dintr-o parte
n alta a mem0ranei alveolo-capilare n unitatea de timp la o anumit di(eren'
de presiune
par'ial.
5apacitatea de di(uziune a plm&nilor pentru >2este n repaus de 8?-2H
ml@min@mm=
dar ea poate crete la CH-DH ml@min@mm= n condi'ii de e(ort sau ,ipo%ie prin
intensi+carea
respira'iei i a circula'iei.
Di(eren'ele alveolo-arteriale ale azelor. :ntre aerul alveolar i s&nele arterial
e%ist i n
mod +zioloic unele di(eren'e privind concentra'ia azelor respiratorii.
<lenusuntimportante
pentru 5>2i 12dar pentru >2radientul poate varia c,iar n repaus ntre ?
mm= la 2H de ani
i 2H mm= la L? ani$ valori care cresc n e(ortul intens.
Acest radient se datorete varia'iei raportului V@" n di(eritele teritorii pulmonare
ca i
contaminrii venoase a s&nelui arterial care atine cam 2R din de0itul cardiac
$%,.transportul 'azelor la nivelul plamanilor
A3
Transportul o%ienului din sinele capilar catre intracelulare de utilizare are loc
printr-un proces de di(uziune.Di(uziunea azelor prin endoteliul capilar si prin
mem0ranele celulare depinde de aceiasi (actori care conditioneaza di(uza-
rea azelor la nivelul plaminului.
"entru a se uni cu o%ienul$ reactie in urma careia se (ormeaza apa$ ionii de
,idroen proveniti din deradarea di(eritelor su0straturi sunt in preala0il activati$
proces care se realizeaza in cursul unui sir de reactii de ozidoreducere ce se
des(asoara in mitocondrii$ cunoscute su0 denumirea de lant respirator./n a0senta
o%ienului reactiile de o%idoreducere ale lantului respirator nu mai au loc si in
consecinta$ este anulata si producerea de enerie$ ceea ce (ace ca procesele
vitale sa nnu mai poata continua
Dio%idul de car0on$care trece din tesuturi in sane$provine din reactiile de
deradare a su0straturilor (reactii de car0onizare) $el reprezentand alaturi de apa
unul din produsii +nali ai reactiilor meta0olice din oranism.
$%8.raportul ventilatie3peruzie eecte asupra pp a 'azelor respiratorii
$%9.centrii nervosi cu rol in re'larea ventilatiei localizati in trunc+iul
cere-ral
Trunc+iul cere-ral parte dorsala -ul- 'rup respirator dorsal care
primeste aerente de la nv I: si :. Eerente pe cai -ul-ospinale la
motoneuronii nn renici 0&70) ale maduvei. 0entrul apneustic ; partea
caudala pontina cu rol minor. <rup respirator dorsal nucleul
pre6otzin'er intre nucle tractului solitar si nucl reticulat -ul-ar, cu
unctie senzitiva,motorie, inte'rare. =euroni cu activitate ritmica in
panta cu rol de pace ma9er> activitatea este determinata de canale de
potasiu de tip A. In e1piratie cand nu descarca, semnale din 'r ventral
intrarzie oprirea activitatii la acest nivel astel incat inspir;%s,
e1pir;&s.
0one1iuni cu alti centri nervosi2
7+ipotalamus termore'lare2ri' prin impulsuri de la termireceptori. ?a
cald creste ventilatia.
7sist li-ic stimul psi+o emotional2spaima oarte mare opreste
inspiratia, emotiile puternice cresc ventilatia.
7scoarta7eerentele catre motoneuronii renici pleaca prin f-re
corticospinale> in control voluntar se poate produce apnee voluntara.
@urata apneei variaza cu sensi-ilitatea c+emoreceptorilor. Apnee
prelun'ita la copii.
7centrul de'lutitiei din tr cere-al, masticatiei, vomei inte'reaza unctia
di'estiva si respiratorie
Arc re/e1 receptorii2 amplasati in ap toraco pulmonar propriu zis si in
aara.ap toraco pulmonar ormat din receptori pulmonari si cai
respiratorii.
$&!.centrii nervosi superiori cu rol in re'larea ventilatiei
5entrii nervoi superiorisitua'i n ,ipotalamus$ sistemul lim0ic i scoar'a cere0ral
realizeaz adaptar-
ea comportamental i voluntar a respira'iei intervenind n termorelare$ strile
a(ectiv-emo'ionale sau
n activit'ile voluntare.
1umeroase re-e%e in-uen'eaz activitatea respiratorie : re-e%ele =erin-Ereuer
i parado%al =ead
ini'iate de distensia sau cola0area plm&nilor$ re-e%ele proprioceptive i
c,emoreceptoare
Activitatea centrilor respiratori 0ul0o-pontini este modulat prin in-uen'a
numeroa-
AL
selor impulsuri provenite din alte teritorii ale sistemului nervos central realiz&nd
adaptarea
comportamental i voluntar a respira'iei n di(erite condi'ii +zioloice.
=ipotalamusulintervine n modi+crile respiratorii leate de termorelare la unele
specii
animale$ n cursul (e0rei sau al unor modi+cri ,emodinamice. Animalele cu
0lan lipsite de
lande sudoripare (c&ine) prezint o ,iperventila'ie intens pentru a evita
supranclzirea.
Ve,icularea unor volume crescute de aer prin cile aeriene superioare produce o
important
pierderedecldurprinevaporare.=ipocapniaesteevitatdeoarecerespira'iaestesup
er+cial
interes&nd doar spa'iul mort.
=ipertermia n cursul (e0rei$ al e(orturilor intense sau e%punerea la temperatur
ridicat
induce ,iperventila'ie prin ac'iunea direct a temperaturii$ accelerarea proceselor
meta0olice
i producerea de meta0oli'i acizi.
=ipotermia are e(ecte inverse deprim&nd respira'ia
$&$.'rupul respirator dorsal
Yrupul dorsal con'ine neuroni cu rol inspirator situat n zona nucleului tractului
solitar care
controleaz activitatea dia(ramului prin intermediul nervilor (renici av&nd rolul
principal n enerarea
ritmului respirator
Yrupul respirator dorsal este situat n zona nucleului tractului solitar i con'ine
mai ales
neuroni inspiratori care controleaz activitatea dia(ramului prin intermediul
nervilor (renici. <i
trimit numeroase +0re spre neuronii rupului respirator ventral de la care pleac
ns mult mai
pu'ine +0re spre rupul dorsal$ ridic&nd semne de ntre0are cu privire la
via0ilitatea mecan-
ismului inerva'iei reciproce n enerarea ritmicit'iispontaneinspira'ie - e%pira'ie.
Se descriu 2 tipuri de neuroni cu rol inspirator: 8
Qcaresuntin,i0a'idein(la'iaplm&nului
i 8 F care sunt e%cita'i de in(la'ie. Yrupul respirator dorsal con'ine i o mic
popula'ie de neu-
roni e%piratori amesteca'i printre cei inspiratori ca i celule " care sunt
interneuroni implica'i
n interarea a(eren'elor de la receptorii pulmonari de ntindere.
1ucleul tractului solitar este sediul proiec'iei +0relor a(erente ale
loso(arinianului (/^)
i vaului (^) care transmit in(orma'ii de la c,emo i 0aroreceptorii carotidieni i
aortici le-
ate at&t de p>2$ p5>2i p= din s&nele arterial c&t i de nivelul presiunii
arteriale. /n plus sunt
transmise i in(orma'ii de la di(eri'i receptori pulmonari$ mai ales cei de distensie.
Se consider c rupul respirator dorsal de'ine rolul principal n enerarea
ritmului respira-
tor ca i n interarea di(eritelor a(eren'e care i pot in-uen'a activitatea pe cale
re-e%.
AD
Trunc+iul cere-ral parte dorsala -ul- 'rup respirator dorsal care
primeste aerente de la nv I: si :. Eerente pe cai -ul-ospinale la
motoneuronii nn renici 0&70) ale maduvei. 0entrul apneustic ; partea
caudala pontina cu rol minor. <rup respirator dorsal nucleul
pre6otzin'er intre nucle tractului solitar si nucl reticulat -ul-ar, cu
unctie senzitiva,motorie, inte'rare. =euroni cu activitate ritmica in
panta cu rol de pace ma9er> activitatea este determinata de canale de
potasiu de tip A. In e1piratie cand nu descarca, semnale din 'r ventral
intrarzie oprirea activitatii la acest nivel astel incat inspir;%s,
e1pir;&s.
$&%.'rupul respirator ventral
Yrupul ventral este situat n zona nucleilor retro(acial$ am0iuu i retroam0iuu i
con'in at&t
neuroni inspiratori c&t i e%piratori
1euronii din nucleul am0iuu sunt mai ales motoneuroni vaali care inerveaz
muc,ii
(arinelui$ larinelui i ai lim0ii +ind implica'i at&t n inspira'ie c&t i n e%pira'ie.
/n nucleul retroam0iuuneuronii inspiratori sunt situa'i mai mult n zona rostral$
n timp
ce neuronii e%piratori sunt situa'i caudal. Mnii dintre neuronii inspiratori trimit
impulsuri spre
muc,ii intercostali e%terni i c,iar spre dia(ram$ n timp ce restul sunt conecta'i
numai cu
al'i neuroni de la nivelul 0ul0ului.
1euronii e%piratori trimit impulsuri spre muc,ii intercostali interni i a0dominali.
1ucleul retro(acial con'ine n special neuroni e%piratori rupa'i n comple%ul
Eotziner
care in,i0 neuronii inspiratori din rupul dorsal ca i unii dintre motoneuronii
(renici. Mul'i
dintre neuronii e%piratori nu sunt activi n eupnee c&nd e%pira'ia are loc pasiv.
/n mod clasic centrii respiratori pontini ar + reprezenta'i de centrul
pneumota%ic i centrul apneustic
<rup respirator ventral pe toata lun'imea -ul-ului.
Are & re'iuni2
$. re'iunea rostrala nc am-i'uu neuroni e1piratori conectati cu 'r
dorsal, modulan' recventa acestora de descarcare.
%. 5ona mediana intermediara neuroni insp>iratori si cu eerente
transmise la I: si : cu rol in 'enerarea ritmului respirator
&. Partea caudala7nucleul retroam-i'uu,intercostali inter,a-d7
neuroni e1piratori cu rol de a controla mm e1piratori accesori
$&&.centrul pneumota1ic
0onectat cu 'rupul respirator dorsal. Activarea sa in+i-a 'r respirator
dorsal, oprind inspirul si permite e1pirul. Stimularea sa determina
cresterea recventei ventilatiei pe seama e1pirului.
@istru'erea le'aturii intre centrii7 respiratie apneustica caracterizata
prin recventa scazuta, amplitudine crescuta, inspir intrerupt din cand
in cand de suspin e1pirator. 0ontroleaza durata inspirutlui Astimul
puternic inspir !,)s, sla- , inspir 7)sB.
@aca se intrerup si aderentele va'ale catre 'rupul dorsal7
apneuzisul;oprirea in inspir a respiratiei Aleziune la nivel 0)B. 0entrul
apneustic este important la caini.
0one1iuni cu alti centri nervosi2
7+ipotalamus termore'lare2ri' prin impulsuri de la termireceptori. ?a
cald creste ventilatia.
AG
7sist li-ic stimul psi+o emotional2spaima oarte mare opreste
inspiratia, emotiile puternice cresc ventilatia.
7scoarta7eerentele catre motoneuronii renici pleaca prin f-re
corticospinale> in control voluntar se poate produce apnee voluntara.
@urata apneei variaza cu sensi-ilitatea c+emoreceptorilor. Apnee
prelun'ita la copii.
7centrul de'lutitiei din tr cere-al, masticatiei, vomei inte'reaza unctia
di'estiva si respiratorie
Arc re/e1 receptorii2 amplasati in ap toraco pulmonar propriu zis si in
aara.ap toraco pulmonar ormat din receptori pulmonari si cai
respiratorii.
$&(.c+emoreceptorii centrali
5,emoreceptorii centrali sunt rsp&ndi'i la di(erite nivele ale sistemului nervos
central
dar mai ales n zona ventro-lateral a 0ul0ului. 5elulele c,emoreceptoare sunt
in-uen'ate de
cantitatea =4din lic,idul ce(alora,idian i cel intersti'ial$ ele ne+ind n contact
direct cu s&nele
arterial de care sunt separate de 0ariera ,emato-ence(alic prin care doar 5>2
di(uzeaz cu uurin'. 5,emoreceptorii centrali nu sunt in-uen'a'i de ,ipo%ie.
5ompozi'ialic,iduluice(alo-
ra,idianestedi(eritdeceaaplasmei$cantitateadeproteine
este cu mult mai mic$ p5>2este ceva mai mare iar p= este mai acid (L.A2)$
sinurul sistem
tamponsemni+cativ+ind0icar0onatul.Datoritacestui(apt,ipercapniavaducelamod
i+cri
mai mari ale =4n lic,idul ce(alora,idian comparativ cu plasma.
5,emoreceptoriicentralisuntimplica'inmen'inerearespira'ieincondi'iiderepausca
i
n rspunsul de lun durat la in,alarea de 5>2$ spre deose0ire de
c,emoreceptorii peri(erici
care intervin n rspunsurile rapide la 5>
In portiunea ventrala a -ul-ului. Prote.ati contra variatiei -ruste din
mediul intern prin -ariera +emato encealica. 8 putin permea-ila la C si
C0O&, dar oarte permea-ile pentri 0O%. 0o% in ?0R in prezenta A0 se
+idrateaza. 0+emoreceptorii centrali care vor stimula 'r respirator
dorsal se a/a in zona e1t ventrlaterala -ul-ara, raeu -ul-ar,locul
coeruleus, nucleu tract solitar.
In acidoza respiratorie ventilatia creste. #odifcari seni-il la 0O%2
sesi-ilitatea lor scade la varstnici, sportivi de perormanta, persoane cu
-oli pulmonare cronice si in somn. 0onsecinte2 apnee in somn sau modul
de a administra o1i'en la persoane cu -oli pulmonare cronice severe2
+ipercapnie cronica, narcoza 0O%.
Cipercapnia stimuleaza c+emoreceptorii centrali la o presiunea mai
mare de ,!mmC' in alveole si san'e arterial. Cipo1ie in S=0
deprimarea c+emoreceptorilor.
$&).c+emoreceptorii perierici
5,emoreceptorii peri(ericisuntsitua'incorpusculii(lomus)dinzonelesino-
carotidian
i aortic i intervin i n relarea circula'iei. /n(orma'iile de la acetia sunt
transmise pe calea
nervilor va i loso(arinian la neuronii respiratori 0ul0ari in-uen'&nd respira'ia.
5orpusculiiauoiria'ie(oarteintens(2HHHml@8HH'esut)+ind(oartesensi0ililascder
ea
CH
p>2$ creterea p5>2$ scderea p=$ reducerea presiunii arteriale$ ,ipertermie sau
creterea
activit'ii simpatice i vasoconstric'ia ce o
nso'ete.Mecanismulprincarereducerea
p>2arterial in-uen'eaz activitatea c,emoreceptorilor este leat de in,i0area
permea0ilit'ii
pentru U4a mem0ranei celulelor c,emosenzitive urmat de depolarizarea
acestora.
/n condi'ii de eupnee cu normo%ie i normocapnie c,emoreceptorii peri(erici
contri0uie
cu 2HR la controlul ventila'iei$ n timp ce ,iperpneea ,ipo%emic depinde n cea
mai mare
msur de controlul peri(eric
la nivelul 'lomusului carotic si crosei aortei. ?a nivelul 'lomusului
'reutatea de % m', vascularizatie -o'ata2 %!!! ml3$!!' consum mare
de o1i'en;9ml3min este sufcient o1i'en dizolvat fzic in plasma, de (!
ori mai mare ca in creier.
E1ista % tipuri de celule 'lomice2
7 de tip I sensi-ile propriu zise ; structura cel enterocromafne din
#@S, secreta catecolamine,dopamina, receptori pentru dopamina
7de tip II;celule de sustinere. inerv simpatica si parasimpata aerent la
nivelul vaselor din zona.
8actorii care stimuleaza zona2 +ipozia, +ipercapnia si crestereaC;
acidoza. In +ipo1ie scade o1i'enul inactic si creste calciul in citosol si se
eli-ereaza dopamina care stimuleaza ventilatia.
0+emoreceptorii e1pusi la o1i'en la o presiunea de )!!mmC' pana la
*! amplitudinea descarcarii este mica. 0+emoreceptorii perierici se
adapteaza 'reu la +ipo1emie si +ipo1ie2 in -oli respiratorii cronice
+iipo1emia creste ventilatia daca se mentine prea mult timp la o1i'en
crescut dispare sin'urul stimul pentru ventilatie7.
0+emoreceptorii raspund la +ipercapnie2 raspuns mult mai mic decat al
celor centrali dar nu prezinta relatie. 0reste ventilata in acidoza
dia-etica si scade alcaloza meta-olica. 0resteri ale temperaturii locale
in eort fzic stimuleaza c+emoreceptorii perierici ce sunt sensi-ili la
variatii ale presiunii partiale de o1i'en.. in anemii cand scade C6 in
into1icatii cu 0O raspunsul este nul. Ramane aceeasi presiune partiala a
o1i'enului. Cipo1ia +ipo1ica de altitudine se activeaza.
$&*.rolul p0O% in controlul ventilatiei
Rolul CO2 este esen'ial i de aceea a i (ost denumit hormon respirator. <l
ac'ioneaz
direct asupra centrilor. creterea presiunii par'iale a 5>2 (hipercapnee)
declaneaz hiperventilaia.
Scderea p5>2 determin rrirea respira'iei i c,iar oprirea ei (apnee).
5reterea p5>2 poate
stimula respira'ia i prin intermediul c,emoceptorilor alveolari i ai zonelor
re-e%oene.
6elarea umoral a respira'iei const n modularea activit'ii centrilor
respiratori prin
ac'iunea azelor respiratorii 5>2 i >2 i varia'iile de p= ale s&nelui i mai
ales ale lic,idului
ce(alora,idian. Aceste su0stan'e in-uen'eaz centrii respiratori ac'ion&nd at&t
direct asupra
C8
neuronilor respectivi c&t i indirect$ prin intermediul c,emoceptorilor din
zonele re-e%oene ale
aparatului cardiovascular.
$&,.rolul pO% in controlul ventilatiei
6elarea umoral a respira'iei const n modularea activit'ii centrilor
respiratori prin
ac'iunea azelor respiratorii 5>2 i >2 i varia'iile de p= ale s&nelui i mai
ales ale lic,idului
ce(alora,idian. Aceste su0stan'e in-uen'eaz centrii respiratori ac'ion&nd at&t
direct asupra
neuronilor respectivi c&t i indirect$ prin intermediul c,emoceptorilor din
zonele re-e%oene ale
aparatului cardiovascular.
Rolul O2 . Scderea p>2 din s&nele arterial e%cit c,emoreceptorii zonelor
re-e%oene
produc&nd ,iperventila'ie. Ac'iunea scderii p>2 direct asupra centrilor
respiratori are e(ecte mai
sla0e.
$&8.rolul p+ ului in controlul ventilatiei
$&9.receptorii pulmonari7rol in controlul ventilatiei
$(!.receptorii e1trapulmonari rol in controlul ventilatiei
$($modifcarea sensi-ilitatii la 0O% a centrilor respiratori
$(%tipuri de ventilatie
$(&.adaptarea ventilatiei in eort fzic
Adaptarea la eort fzic poate f ventilatorie, circulatorie sau tisulara.
@e-itul de o1i'en creste de la %)! ml in repaus pana la *!!! ml3min.
0reste de ( ori nivelul de rapaus. 0reste recventa cardiaca si
mo-iliarea san'elui din depozit. In eort &)ml3min aport de o1i'en
spre tesut.
?imitarea eortului este acuta de capacitatea tesutului de a se adapta
la eort2 viteza si nr de cap suplimentare desc+ise, distanta mai mica
de B% de deplasare, capacit mitocondriilor de a utiliza O%.
@pdv respirator ventilaia se adapteaza in unctie de eort
moderat,mediu, intens anaero-.
Inainte de de-utul eortului creste ventilatia pe seama amplitudinii apoi
are loc cresterea treptata, lenta pana la var> la oprirea eortului
ventilatia scade -rusc.
?a e1ercitiu mediu tamponare izocapnica a 0o% ventilatie sufcienta
pentru a reduce 0o%.
Eort anaero- acumulare de acid lactic se produce acidoza
o-oseala musculara datorita -om-ardarii motoneuronilor cu impuls de
la mm si tendoane, scade 'lico'enul.
Acumularea de acid lactic determina in/amatie locala a musc+iului si
e-ra musculara.
Particularitati ale respiratiei2
7 tusea mec re/e1 de aparare2e1pir ortat cu 'lota inc+isa, presiune
intrapleurala crescuta pana la $!!mmC'> 'lota se desc+ide -rusc
aerul e1pulzat cu viteza;$!!9m3+7antreneaza particule straine.
7stranut7e1pier ortat cu 'lota desc+isa
7su'+itul7miscarea spasmotica2 in inspir inc+idere -rusca a 'lotei>
oprit prin apnee cateva secunde acidoza , cedarea contr musc.
C2
7cascat7misc insp ampla cu semnifcatie necunoscuta alveolele
cola-eaza in inspir, scade o1i'enarea cere-rala, creste nevoia de
o1i'en.
7respiratia periodica7de tip c+eDen sto9es2 uneori in somn, spontan la
persoane ara -oli respiratorii> det de variatii ale sensi-
c+emoreceptorilor centrali la 0O%> misc ventilatorii cu amplitudine
crescuta, apoi scade si duce la apnee. Apare la persoane cu insufcienta
cardiaca, uremie, aectiuni cere-rale. 0reste timpul de circulatie si
e1tractia de o1i'en, acumulare de co%.
<(ortul +zic presupune creterea aportului de >2pentru a (ace (a' necesit'ilor
meta0olice
crescute nso'it de producerea i eliminarea unei cantit'i sporite de 5>2$
(enomene realizate
prin modi+cri respiratorii adecvate.
5onsumul de >2n repaus (MV>2) este n Bur de 2?H ml@min$ dar poate crete n
e(ortul
intens la A-C 9@min (C?-?H ml@S corp). Valorile sunt ceva mai mici la (emei i scad
cu v&rsta.
/n cursul e(ortului$ n (unc'ie de intensitatea acestuia$ consumul de >atine un
nivel de
ec,ili0ru (steadT state) care se men'ine constant. Atinerea acestui nivel nu se
(ace 0rusc ci
treptat$ iar la terminarea e(ortului consumul revine la normal tot treptat (#/Y
2.2?).
De+citul de >2realizat la nceput va + recuperat su0 (orma datoriei de >2dup
terminarea
e(ortului$datorie care este mai mare dec&t de+citul at&t datorit nereveniriii
mediate a respira'iei$
circula'iei i temperaturii c&t i reo%idrii lactatului produs de activitatea
muscular.
Mn su0iect neantrenat poate acumula o datorie de>
2de p&n la 8H 9$ n timp ce la antrena'iea atine 8L-8D 9.
/n condi'iile respira'iei de aer atmos(eric cu un con'inut normal de >2(28R)
consumul
acestuia crete paralel cu intensitatea e(ortului p&n la atinerea unei limite care
constituie
capacitatea aero0ic ma%im. <a limiteaz capacitatea ma%imal de e(ort a unui
individ care
depinde nu numai de sistemul ventilator ci i de cel cardio-vascular
Adaptarea ventila'iei la e(ortse(aceprin,iperventila'iecareatine82H-
8?H9@minsauc,iar
mai mult la cei antrena'i. =iperventila'ia se realizeaz prin creterea (recven'ei i
amplitudinii
respira'iilor ntre ele sta0ilindu-se o interrela'ie optim n (unc'ie de intensitatea
e(ortului. 9a
nceputcretemaimultvolumulcurent$darpemsurceapareacidozameta0olicpred
omin
creterea (recven'ei.
9ucrul mecanic ventilator crete pe msura intensi+crii e(ortului.
=iperpneea i amploarea ei presupune interven'ia mai multor (actori care lua'i
+ecare n
parte nu au e(ectul sumrii lor$ e%ist&nd importante varia'ii individuale.
Adaptareaventila'ieiarelocprinmecanismec,emoneuroloicecomple%encincompl
et
CA
cunoscute.
Mecanismele neuroene sunt leate de:
-in-uen'ele peri(erice de la nivelul articula'iilor i muc,ilor a-a'i n activitate ca
i de la
c,emoreceptori
-in-uen'ele centrale prin creterea temperaturii i a p5>
$((.ventilatia in conditii de +ipo-arism
6espira'ia n ,ipo0arism. =ipo0arismul se caracterizeaz prin scderea p> pe
msur ce altitudinea crete supun&nd oranismul unei ,ipo%ii mai mult sau mai
pu'in accentuat
<(ectele sale depind de nivelul altitudinii$ rapiditatea cu care suntem e%pui i
radul de
e(ort.
<volu'ia semnelor clinice dei este varia0il n (unc'ie de individ se (ace n mai
multe
etape:
-(aza indi(erent (p&n la AHHH m) (r modi+cri importante n a(ara unei
adaptri pre-
lunite la ntuneric.
-(aza de ,ipo%ie com-pensat(AHHH-C?HH m) se caracterizeaz prin intensi-
+careaventila'iei$
creterea (recven'ei cardiace$ de+cien' uoar n realizarea unor acte
psi,omotorii comple%e$
scderea memoriei i a capacit'ii ma%imale de e(ort.
-(aza de ,ipo%ie mani(est (C?HH-3HHH m) provoac deradarea proceselor
mentale i a
controlului neuro-muscular$ pierderea capacit'ii de Budecat critic$ letarie$
indi(eren' sau
e%cita'ie eu(oric$ ,alucina'ii. Ventila'a i (recven'a cardiac cresc n continuare.
-(aza de ,ipo%ie critic (3HHH-DHHH m) determin o deteriorare rapid a Budec'ii
i
n'eleerii$ pierderea controlului neuro-muscular i a contien'ei$ oprirea
respira'iei$ a circula'iei
i moartea.
Semnele ,ipo%iei severe nso'ite de tul0urri nervoase devin aparente la om c&nd
p>
scade su0 AH mm= la nivel alveolar. Acest nivel este ns mult mai mare dac
,ipo%ia se
com0in cu ,iperventila'ie i ,ipocapnie deoarece circula'ia cere0ral este (oarte
sensi0il
la varia'iile p5>
Deconectarea 0rusc a alimentrii cu >la altitudini mari duce rapid la com (A
min la
GHHH m i mai pu'in de 8 min la peste 82HHH m).
9a nl'imi mari tul0urrile nu pot + prevenite nici prin administrarea de >8HHR
dac
presiunea acestuia nu este su+cient pentru a crete presiunea sa par'ial la
nivel alveolar.
Aceasta datorit (aptului c presiunea vaporilor de ap +ind independent de
altitudine la
temperatura corpului$ volumul ocupat de aceti vapori crete pe msura scderii
presiunii
atmos(erice.
CC
9a 8?HHH m suma p5>i a vaporilor de ap ealizeaz pe cea atmos(eric i
plm&nii nu
vor mai con'ine > iar la 8GHHH m presiunea atmos(eric este ealat de cea a
vaporilor de
ap (CL mm=) care vor ocupa ast(el ntre plm&nul
=ipo0arismul acut provoac modi+cri la nivelul ventila'iei pulmonare$ a (unc'iei
respiratorii a s&nelui i a aparatului cardio-vascular.
Ventila'ia pulmonar crete ca urmare a stimulrii ,ipo%ice a c,emoreceptorilor
peri(erici. 5,emoreceptorii centrali nu reac'ioneaz la ,ipo%ie$activitatea lor +ind
c,iar deprimat din cauza ,ipocapniei i alcalozei la nivelul lic,idului ce(alo-
ra,idian
Aclimatizarea se realizeaz prin:
-creterea ventila'iei alveolare ca urmare a impulsurilor c,emoreceptoare prin
scderea
p>
-aBustareaec,ili0ruluiacido-
0azicals&neluiilic,idelororanismuluiprincontracararea
alcalozei respiratorii produs prin ,iperventila'ie ca urmare a scderii 0azelor
-creterea capacit'ii de transport a >
de ctre s&ne prin polilo0ulie i prin scderea
a+nit'ii pentru >a ,emolo0inei ca urmare a creterii 2-A D#Y cauzat de
alcaloz i (avoriza-
rea cedrii >la 'esuturi
-creterea de0itului cardiac i redistri0u'ia s&nelui n primele (aze ale e%punerii
-creterea numrului de capilare musculare i a nivelului enzimelor o%idative
celulare
5a urmare a acestor mecanisme locuitorii marilor nl'imi pot e(ectua e(orturi mai
mari$
consum mai pu'in >pentru aceste e(orturi$ respir mai mult aer pentru a o0'ine
> necesar
i au modi+cri mai mici ale (recven'ei cardiace i presiunii arteriale.
Aclimatizarea este un proces reversi0il de ndat ce condi'iile mediului se
sc,im0.
9ipsa aclimatizrii datorit incapacit'ii c,emoreceptorilorde a in-uen'a respira'ia
duce la
apari'ia N0olii cronice a nl'imilorOcaracterizat prin ce(alee$o0oseal$dureri ale
e%tremit'ilor$
tul0urri vizuale i de contien'
$().ventilatia in conditii de +iper-arism
6espira'ia n ,iper0arism. Scu(undarea su0acvatic (sca(andrii$ c,esonieri$
scu(undtori autonomi) in-uen'eaz n mod semni+cativ respira'ia datorit
(aptului c la
+ecare 8H m presiunea crete cu 8 atm.
Scu(undarea n apnee i revenirea la supra(a' n scurt timp este posi0il (r nici
un
ec,ipament$ncondi'iie%perimentalerealiz&ndu-
sescu(undrilaad&ncimidepeste2HHm./n
urmaantrenamentuluiscu(undtoriiprezintcreterea5"T$diminuareaV6$toleran'c
rescut
la 5>2i la ,ipo%ie.
5,iar scu(undarea corpului n ap p&n la &t are e(ecte asupra respira'iei i
circula'iei
C?
dei presiunea e%ercitat asupra toracelui nu depete presiunea atmos(eric cu
mai mult
de 2H cm ap. Are loc o scdere a 56# pe seama V<6 din cauza creterii
volumului sanuin
pulmonar odat ce ntoarcerea venoas este mrit de presiunea e%ercitat din
a(ar asupra
veneloriavenoconstric'ieisimpaticedactemperaturaapeiestemaisczut.9ucrulm
ecanic
respirator crete.
/mersia capului sau numai a (e'ei poate produce o 0radicardie accentuat nso'it
de
creterea presiunii arteriale sistemice. Acest re-e% de Nscu(undareO este vizi0il
mai ales c&nd
apa este rece sau la animalele i psrile acvatice.
"entru a putea respira n cursul scu(undrilor de mare ad&ncime i lun durat
presiunea aerului respirat tre0uie s +e eal cu cea ,idrostatic.
Volumul azelor la nivelul plm&nilor scade odat cu creterea presiunii ceea ce
reduce n mod corespunztor 5"T.Ast(el scu(undareala2Hm unde presiunea atine
A atm reduce volumul plm&nilorla8@A din volumul de la supra(a'.
6espira'ia su0acvatic este posi0il doar cu aButorul unor ec,ipamente speciale
care asiur o presiune a aerului respirat corespunztoare
ad&ncimii de scu(undare
/n scu(undrile de mare ad&ncime ,ipercapnia la care se asociaz ac'iunea
narcotic a
azotului dizolvat n s&ne ntr-o cantitate mai mare con(orm presiunii sale par'iale
duce la aa
numita N0e'ie a ad&ncurilorO.
<(ectele creterii concentra'iei 5>2 depind de nivelul acestuia$ timpul de
e%punere i
caracteristicile individuale. "ersoanele care tolereaz mai 0ine ,ipercapnia au o
respira'ie mai
lent iar V5$ V/6 i 5V sunt mai mari.
=ipercapnia acut produce ncep&nd de la concentra'ii de 8.? R ,iperventila'ie$
ta,icar-
die$ ,iperlicemie datorit creterii tonusului simpatic. /n cazul e%punerilor
cronice alterarea
(unc'iilor +zioloice se mani(est la concentra'ii de peste A R.
Dis0arismul. /n timpul scu(undrilor$ din cauza creterii presiunii par'iale a
azelor$ azotul
care este un az inert se dizolv n s&ne i 'esuturi n cantit'i mai mari.
Ascensiunea 0rusc
duce la eli0erarea azotului su0 (orm de 0ule a cror dimensiune crete pe
msura ascensiunii
duc&nd la importante tul0urri caracterizate prin dureri articulare$ ta,ipnee$ tuse$
(urnicturi
cutanate$ tul0urri de vedere$ ce(alee$ dureri a0dominale.
Mneori predomin tul0urrile neuroloice (paralizii$ convulsii$ com)$ alteori cele
cardio-
vasculare produse datorit ,emoconcentra'iei i a creterii presiunii arteriale.
Situa'ii similare$ dar mai pu'in rave$ se pot nt&lni la aviatori n cazurile
ascensiunii (oarte
rapide sau a decomprimrii 0rute a ca0inelor presurizate
<(ectele to%ice asupra sistemului nervos sunt o0servate la presiuni mai mari de 2
atm i
C3
se mani(est prin tremurturi$ convulsii i pierderea contien'ei. <le se nso'esc
de tul0urri
de vedere i ale (unc'iei renale. Acestea apar cu at&t mai repede cu c&t presiunea
i durata
e%punerii sunt mai mari$ mai ales n condi'ii de e(ort.
Administrarea de 5>nrut'ete situa'ia at&t prin ac'iunea sa direct c&t i prin
e(ectele
acidozei.
<m0olia azoas care nu tre0uie con(undat cu dis0arismul poate apare n cazul
n care
unscu(undtorcarerespiraerlaopresiuneridicatieserapidlasupra(a'(ramairesp
ira./n
aceste condi'ii volumul aerului din plm&ni crete 0rusc i poate cauza ruperea
vaselor pulmo-
nare cu ptrunderea aerului n sistemul circulator i apari'ia em0oliei azoase.
#enomenul nu
apare n cazul celor care respir aerul de la supra(a' se scu(und i apoi revin la
supra(a'.
$(*.tipuri de +ipo1ie
Cipo1ia +ipo1ica; scadere a presiunii partiale de o1i'en2 in conditii de
altitudine sau cand este aectata diuziunea alveolo capilara sau sunt
dr7st' cu cianoza.
Cipo1ia anemica7 scade nr de transportori pt o1i'en2 in anemnii cand
numarul de +ematii si +- scad, dar si in into1icatii cu co%.
Cipo1ia sta'nanta2 aecteaza tesutul la nivelul caruia se maniesta
e1punere la ri', insufcienta cardiaca.
Cipo1ia +istoto1ica7 in into1icatii cu cianuri
C.Cipo1ica la e1punere -rusca de altitudini crecute la &!!!m
presiunea alveolara a !%;*!mmC'7pctul critic la care c+emoreceptorii
perierici sunt stimulati. &,!!;primele simptome de +ipo1ie2
irita-ilitate,insomnie,inapetenta. ))!!7 simptome nervoase
e-rietate,conuzie,ameteli,'reata,varsaturi,incapacitate de orientare
temporo spatiala.instalarea edemului cere-ral si pulmonar cu coma si
deces. $9!!!m,;pres atmoserica;(,mmC'7presiunea vap de C%O din
or'anism.
Adaptarea la altitudine2
7acuta cresterea ventilatiei2 +ipo1ia +iperventilatia determina
eliminarea crescuta de co%;alcaloza care deplaseaza la stan'a cur-a C-
7cronica, dupa 8 zile2eritropoietina determina cresterea numarului de
eritrocite, nr de mitocondrii tisulare si capacitatii de e1tractie a
o1i'enului. ?ocalnicii au poli'lo-ulie, sunt cianotici, torace in -utoi.
Riscurile e1punerii -ruste la altitudine2 edem cere-ral2 rie
7raspuns motor cere-ral la +ipo1ie;vasodilatatatie
7edem pulmonar2 +ipo1ia determina+ipertensiune pulmonara cu
vasoconstrictie.
Persoanele cu diureza crescuta se adapteaza cel mai -ine7 se eli-ereaza
ad+, creste volemia, presiunea arteriale edem cere-ral si pulmonar.
Tratament cu acetazolamida care in+i-a secretia de potasiu si
stimuleaza secretia natriului.cortizolul scade permea-ilitatea capilara.
In+i-itori de canale de calciu2 niedipina.
$(,.+ipo1ia anemica
7 anemii2 in repaus eectele respiratorii nu sunt severe decat la anemii
ma.ore2 p)!crescut, C- cedeaza mult O%
CL
7into1icatia cu 0O2 C- scade valoarea lui p)!> +- cedeaza 'reul la !%>
este letala cand ,!E din +- este -locata.
Terapia cu o1i'en $!!E 7 ma1im (7) zile la persoane cu -oli cardio
con'enitale, -oli pulmonare cronice. Poate determina iritatia cailor
respiratorii, scaderea activitatii cililor din cai resp superfciale, tuse
spastica, leziuni ale tractelor resp> la nou nascuti prematori viroza
c+istica pulmonara si f-roplazie retroletala;de'enerarea vaselor
retiniene datorita an'iotensinei la e1punerea la !% in e1ces.
$(8.to1icitatea o1i'enului
Dei o%ienul molecular este indispensa0il pentru viata$ in acelasi timp el are un
e(ect neativ$ cunoscut ca to%icitatea o%ienului$ determinata de (ormarea unor
specii reactive
To%icitatea o%ienului se mani(est la nivelul sistemului nervos central. "resiuni
par'iale ale o%ienului cuprinse ntre H$28 0ar (sc. a0s.) i H$C2 0ar (sc. a0s.) se
consider a + normale (normoo%ic). =ipero%ic este un amestec de az (aer
comprimat sau amestec respirator sintetic) la care presiunea par'ial a o%ienului
din este mai mare de H$C2 0ar (sc. a0s.).
=ipero%ia poate + cronic sau acut. =ipero%ia apare datorit o%ienului con'inut
de aerul comprimat respirat la o presiune par'ial ridicat av&nd un e(ect to%ic
asupra oranismului.
=ipero%ia cronic (e(ectul 9orrain Smit,) apare atunci c&nd presiunea par'ial a
o%ienului din amestecul respirator este cuprins ntre H$C2 0ar (sc. a0s.) i 8$L
0ar (sc. a0s.). =ipero%ia cronic este 0ine tolerat de ctre sca(andri n cursul
unor scu(undri de scurt durat.
=ipero%ia acut (e(ectul "aul Eert) sau criza ,ipero%ic apare atunci c&nd
presiunea par'ial a o%ienului din amestecul respirator este mai mare de 8$L 0ar
(sc. a0s.). =ipero%ia acut tre0uie evitat deoarece poate avea consecin'e rave
Toate oranismele sunt e%puse la 0oli si sunt puternic adaptate la conditiile
ne(avora0ile. /n acest conte%t$ plantele$ animalele si oranismele mici sunt
inrudite intre ele. Atat 0olile cat si adaptarile la mediul inconBurator sunt leate
(recvent de alterarea meta0olismului c,imic al o%ienului caracterizat prin
tranzitia de la ,eterolitic (2 electroni atrasi sau cedati) pana la cresterea
proceselor ,omolitice (8 electron tranzitat). 6eactiile ,omolitice produc radicali
care pot reactiona in mod aresiv deteriorand compartimente celulare$ tesuturi si
oranisme. De aceea$ aceste reactii enerand specii reactive ale o%ienului sunt
contra0alansate$ in eneral$ de cresterea interna a proceselor o%idative sau de
aButorul medicamentos e%tern
$(9.comportamentul +- ata de o1i'en
$)!.P)! O%
$)$.cur-a de asociere disociere a C-
5ur0a de disociere a o%i,emolo0inei. #i%area >2de ,emolo0in se (ace la
nivelul
plm&nilor$ iar des(acerea sa la nivel tisular n condi'iile varia'iei presiunii par'iale
a >
2ca i a altor (actori. Aceste procese au loc prin modi+cri ale con+ura'iei
moleculei de ,emolo0in
i constituie aa numita respira'ie molecular.
6eprezentarea ra+c a interrela'iei dintre satura'ia ,emolo0inei i p>2arat ca
o cur0 n (orm de S ce caracterizeaz enzimele relatoare
CD
#orma aceasta e%plic de ce prima molecul de >2 se +%eaz i se cedeaz mai
reu.
Satura'ia ,emolo0inei$ c,iar la p>2 de 8HH mm=$ nu este 8HHR ci doar G?-
GDR din cauza
contaminrii s&nelui arterial cu s&ne venos prin leturile dintre circula'ia
pulmonar i cea
0ronic ca i prin vrsarea s&nelui prin unele vene coronare n ventriculul
st&n.
>seriede(actoriin-uen'eazcur0adedisociereao%i,emolo0ineipecareodeplaseaz
la st&na$ ceea ce reprezint o capacitate de +%are crescut a >2$sau spre
dreapta nsemn&nd
o capacitate de cedare crescut
$)%.rolul p+ ului asupra relatiei C-7O%
6olul ,emolo0inei n men'inerea p=. =emolo0ina ca orice protein are un
caracter
am(oter ceea ce (ace ca n mediul alcalin al s&nelui (p= L.C) s se ncarce
neativ i s se
comporte ca un acid sla0 le&nd 0aze. 5um ncrcarea electric depinde de p=
punctului izo-
electric$ o%i,emolo0ina este un acid mai puternic (p= izoelectric la 3.3) dec&t
,emolo0ina
redus (p= 3.D).
/nacestecondi'iitrecereaunuimoldeo%i,emolo0inla,emolo0inredusva+nso'it
de learea a H.3-H.L m<W =4ceea ce aBut la men'inerea ec,ili0rului acido-0azic.
#enomenele au loc invers la nivelul plm&nilor iar =4eli0erat va reac'iona cu
=5>A-d&nd natere la =5> A = care disociaz la 5>2i ap
5antitatea mare de ,emolo0in din s&ne (ace ca rolul su de tamponare s +e
(oarte
important.
$)&.rolul 0O% asupra relatiei C-7O%
6olul 5>2i al =4n modi+crile respiratorii este (oarte important$ lucru dovedit
de
creterea aproape liniar a ventila'iei n (unc'ie de concentra'ia =4. 1ivelul
acesteia depinde
de producerea meta0oli'ilor acizi i n primul r&nd a 5>2 care este cel mai
puternic e%citant al
neuronilor respiratori. =ipo%ia poten'eaz rspunsul ventilator la 5>2.
"resiunea par'ial a 5>2n s&nele arterial i lic,idul ce(alo-ra,idian constituie
principala
determinant a volumului curent i a (recven'ei respiratorii. Doar ,iperventila'ia
voluntar i
,iperpneea din cursul e(ortului pot depi e(ectul ,ipercapniei asupra respira'iei.
<(ectele cele mai marcate ale 5>2se o0'in la concentra'ii de?-8HR n aerul
respirat nso'ite
de varia'ii ale p5>2 arterial ntre CH-LH mm=. Depirea acestor concentra'ii nu
are e(ecte
vizi0il mai marcate$ dar creterile peste LH-DH mm= au e(ecte in,i0itoare asupra
respira'iei
nso'indu-se de dispnee$ ce(alee intens datorit vasodilata'iei cere0rale$ stare
de nelinite$
ame'eli sau lein. "ierderea contien'ei$ riiditatea muscular i tremorul apar la
concentra'ii
de 5>2ce depesc 8?R. "este 2H-AHR se produc convulsii eneralizate
CG
Deprimarea rspunsului la 5>2p&n la apnee poate apare n somn$ dup
consumul de
su0stan'e opioide$ 0ar0iturice sau anestezice put&nd cauza moartea. A(ec'iunile
pulmonare
cronice o0structive deprim rspunsul ventilator la ,ipercapnie at&t datorit
dezec,ili0rului
acido-0azic c&t i din cauz c lucrul mecanic ventilator crescut nu permite
realizarea unei
,iperventila'ii corespunztoare e(ortului
$)(.rolul %,& 6P< asupra relatiei C-7O%
scade afnitatea pentru O
%
a C- cedarea O
%
la Fesuturi.
7 mecanism2 se f1eaza ec+imolecular pe lanFurile - ale C-A A
%
%
B
modifcGri conormaFionale cu eli-erarea O
%
de pe C-O%.
7 deplaseazG cur-a de disociere a C-O
%
la dreapta, mGrind eli-erarea O
%
la nivel tisular
$)).enomenul 6o+r
$)*.enomenul +am-ur'er la tesuturi
0o%7 diuzeaza din tesuturi in plasma 'radient de presiune ()7(*
mmC', in tesut la (! mmC' in san'e arterial7 O% diuzeaza din san'e in
tesuturi conorm 'radientului presional 9)79, la(!mmC'7
,E din co% este dizolvat in plasma
0o diuzeaza in +ematie unde se com-ina cu +%!
Cco& diuzeaza in plasma la sc+im- cu cl care diuzeaza in +ematie si
impreuna cu na ormeaza un sistem tampon. Iar ,!E din co% este
transportat su- orma de sist tampon =aC0o&.
Restul de %&E din co%, se transporta ca si car-amin +emo'lo-ina
$),.orme de transport a 0O% in san'e
Dio%idul de car0on (ormat la nivelul tesuturilor este transportat in di(erite
moduri.
- dizolvat in plasma$ ca si in cazul o%ienului$ o parte din dio%idul de car0on$
care di(uzeaza dinspre tesuturi prin lic,idul interstitial$ in sine este transportat
dizolvat in plasma.
- leat de anumite rupuri ale anumitor proteine o cantitate de apro%imativ
de dio%idd de car0on la 8HHml.sine se leaa la nivel unor rupari ale proteinelor
plasmatice$ inclusiv la nivelul ,emolo0inei$ +ind transportat su0 aceasta (orma.
- su0 (orma de 0icar0onat. su0 aceasta (orma se transporta restul de dio%id
de car0on din sine.Dio%idul de car0on di(uzat de la nivelul tesuturilor in plasma$
patrunde in interiorul eritrocitelor unde su0 in-uienta unor enzime an,idraza
car0onica$ se ,idrateaza dind nastere acidului car0onic. Acidul car0onic
disociaza rapid iar anionul 0icar0onic rezultat di(uzeaza din nou$ in cea mai mare
parte in plasma unde leaa ionul 1a pozitiv cu care (ormeaza 0icar0onatul de
sodiu.
Transportul 5>2se (ace at&t n plasm c&t i n ,ematii. 5on'inutul n 5>2 al
s&nelui arterial este de CD-?H volume R iar al celui venos de ?2-?D volume
R.5>2are o solu0ilitate mai mare dec&t >2 ast(el nc&t n plasma din s&nele
arterial unde p5>2este de CH mm= s avem dizolvat o cantitate de 2.G ml@8HH
ml s&ne (L-DR).
6estul de 5>2se sete at&t n plasm c&t i n ,ematii su0 A (orme :
-o mic parte su0 (orm de =5>A=
-cea mai mare parte (DHR) su0 (orm de 0icar0ona'i (=5>A1a i =5>AU)
-su0 (orm car0aminic leat de proteinele plasmatice sau mai ales de
,emolo0in
(?-8HR
?H
Dei (orma car0aminic nu este n cantitate prea mare ea este (oarte mo0il ceea
ce (ace
ca n Bur de 8@C din sc,im0urile de 5>2s se realizeze prin intermediul acestei
(orme
5onsumul de >
2
depindedenecesit'ileenereticeale'esuturilorn(unc'iedeactivitatea
depus. 5antitatea de 5>2(ormat i eliminat variaz n (unc'ie de modi+crile
meta0olice$
respiratorii i circulatorii
8. #orma dizolvat +zic a 5>2 (?R)
K partea di(uzi0il - determin sensul i mrimea di(uziunii$
+%area su0 (orm de car0amat@0icar0onat
2. #orma com0inat cu proteinele plasmatice i ,emolo0ina (C$?R).
5>2 +%at de ruprile aminice ale:
- proteinelor plasmatice car0ama'i
- =0 car0=0
A. 5>2 transportat su0 (orm de 0icar0onat (GHR)
- U=5>A intraeritrocitar si 1a=5>A n plasm.
:n plasm: o cantitate 5>2 se ,idrateaz spontan =25>A =5>A- 4 =4
5ontrolul (ormrii i eli0errii 5>2 de pe =0
'radul de o1i'enare al C- Aeect CaldaneB2 O
%
tinde
sG eli-ereze 0O
%
.
7 la Fesuturi2 eli-. O
%
de pe C- avorizeazG f1area 0O
%
7 la plGmHni2 O
%
determinG eli-erarea 0O
%
din C-0O
%
$)8.presiunile partiale ale 'azelor in tesuturi,plamani si san'e
$)9.orme de transport a O% in san'e
>%ienul este transportat in sane su0 doua (orme :
- dizolvat in plasma. desi in cantitatea mica$o%ienul transportat su00
aceasta (orma are un rol (unctional deose0it$ reprezentind(orma intermediara
o0liatorie in trans(erul de o%ien intre aerul alveolar si ,emolo0ina din eritrocit
sau intre ,emolo0ina eritrocitara si celulele catre care o%ienul este eli0erat.
- leat de ,emolo0ina. reprezinta (orma principala de transport a
o%ienului$ su0 aceasta (orma sunt transportati apro%imativ 2H ml. o%ien in
+ecare ml. de plasma$ +ecare ram de ,emolo0ina leind 8$AC ml. o%ien. 9a
+ecare atom de #e din structura moleculei de ,emolo0ina se leaa la0il cite o
molecula de o%ien.
5apacitatea ,emolo0inei de a lea o%ienul este conditionata nu numai de
presiunea partiala a o%ienului dizolvat ci si de concentratia ionilor de ,idroen
din plasma si de temperatura.5resterea temperaturii si a concentratiei ionilor de
,idroen scade capacitatea ,emolo0inei de a lea o%ienul care este cedat
tesuturilor
Transportul >2se realizeaz n cea mai mare parte de ctre ,emolo0in.
Aceasta poate lea 8.AC ml >2pe +ecare ram ceea ce corespunde la o valoare n
Bur de 2H
ml@8HH ml s&ne.> mic parte a >2(cam H.A ml R) se dizolv n plasm con(orm
leilor +zicale. Aceast (orm este important pentru c prin ea au loc toate
sc,im0urile de la nivelul ,ematiilor spre 'esuturi sau de la plm&ni spre ,ematii.
Viteza de reac'ie dintre >2i ,emolo0in e%primat
prin atinerea satura'iei de ?HR este (oarte mare permi'&nd
casc,im0urilesse(acncondi'ii
optime c,iar la o vitez crescut de curere a s&nelui
5onsumul de >2depinde de necesit'ile eneretice ale 'esuturilor n (unc'ie de
activitatea
?8
depus. 5antitatea de 5>2(ormat i eliminat variaz n (unc'ie de modi+crile
meta0olice$
respiratorii i circulatorii
SA=<E
83H.=ematocritul
Hematocritul reprezinta volumul de globule rosii raportat la volumul de sange, exprimat in procente. Determinarea Ht se
#ace pe singe venos recoltat pe anticoagulant in tuburi capilare
cu pereti grosi' prin centri#ugare' la 8000turatii/min. Hematiile' cu densitatea de 1090
sedimenteaza la baza tubului' iar plasma deasupra coloanei celulare. La limita de separare
dintre ele se detaseaza un strat albicios de lim#o1monocite si trombocite' a caror densitate
este intre cea a +ematiilor si cea a plasmei. !entru determinarea corecta a Ht' este
necesara aplicarea unor #actori de corectie ce tin de anticoaglantele #olosite si de plasma
sec+estrata in urma centri#ugarii intre elementele #igurate din singele venos #olosit. Ht. venos
de 5& este putin mai mare #ata de cel arterial' 2&' datorita trans#erurilor +idroelectrolitice
de la nivelul segmentului capilar= +ematia din singele venos contine mai mult clor si mai
multa apa datorita #enomenului de
membrana Hamburger.Baloarea Ht variaza si in #unctie de organul in care este determinat= la
nivel splenic' Ht este de 70&' la nivelul capilarelor sinusoidale care E#iltreazaF eritrocitele. La
nivelul capilarelor si venulelor' Ht este mai mic' datorita curgerii a$iale a singelui' iar la
nivel tisular' este mai mic #ata de Ht arterial si venos. Ht scade in anemii'
cu volemie normala sau scazuta' in +iper+idratari )+idremii* situatie in care volumul
globular este normal' dar Ht scade datorita +emodilutiei. 3n cazul +emoragiilor acute' inaintea
declansarii mecanismelor compensatorii pentru corectarea +ipovolemiei prin intravazarea
lic+idului interstitial' Ht este normal' dar volumul globular este scazut.
$*$."ariatii fziolo'ice ale volemiei
Digestia determina o usoara si lenta crestere a volemiei' datorita absorbtiei lic+idelor in
intestin.
&n timpul efortului fi)ic intens' volemia scade cu citeva sute de mL in primele 10115
min.' datorita e$travazarii de lic+id in spatiul interstitial' datorita cresterii nr. de capilare
#unctionale. La subiectii antrenati' acest proces e mai putin intens. Bolemia este direct
proportionala cu activitatea #izica= la sportivii bine antrenati' raportul volum sanguin/2
corp este de 100 mL/Igc.
PosturaA dupa 80 min de ortostatism' volumul sanguin este cu 15& mai mic #ata de
clinostatism' datorita e$travazarii lic+idului din plasma la nivelul capilarelor membrelor
in#erioare' ca urmare a cresterii presiunii +idrostatice la acest nivel.
#epaosul la pat pe termen scurt determina cresterea volemiei cu 5&. !relungit' )218
saptamini* acesta determina scaderea volumului plasmatic la adult cu pina la 500 mL.
Sarcina determina cresteri mari ale volemiei' in medie cu 20180&' uneori cu 100&.
7resterea volemiei incepe din saptamina 10 si continua progresiv pina in saptamina 801
8' raminind nemodi#icata pina la nastere. 0evenirea la normal se #ace lent' dupa 21% sapt
de la nastere. 5ceasta crestere se datoreaza cresterii atit a volumului plasmatic )B!*' cit si
a volumului globular )B2*. B! creste datorita retentiei +idrosaline din timpul sarcinii' ca
urmare a secretiei crescute de aldosteron si 5DH' si datorita cresterii sintezei de proteine
plasmatice. 7resterea B2 in sarcina variaza direct proportional cu aportul de Fe e$ogen.
7auza cresterii de B2 este descarcarea de eritropoietina' stimulata de prolactina si de
Hormonul lactogen placentar )somato1mamotropina corionica umana' denumita si
prolactina placentara " +L!*. Factori e8ogeni. "limatul cald induce usoara crestere a
volemiei' iar cel rece o usoara scadere' datorita trans#erului de lic+id in tesuturi )#icat'
musc+i*. La mare altitudine' presiunea atmos#erica scazuta si +ipo$ia prelungita determina
cresterea volemiei.
$*%.Cipovolemiile normocitemice,oli'ocitemice si policitemice
?2
(caderea volumului sanguin total reprezinta
hipovolemiile sau oligohemiile. 5cestea apar in= pierderi e$terne de singe )traumatisme'
interventii c+irurgicale* sau interne )ruptura de organ= #icat' splina*. (caderea volumului
globular caracterizeaza anemiile. (caderea B! se datoreaza pierderii de plasma si
lic+idelor +idroelectrolitice in arsuri' varsaturi' diarei pro#uze' #istule digestive' ocluzii
intestinale inalte' diureze e$cesive' diabet za+arat )poliurie*' insu#icienta 70' transpiratii
pro#uze' edeme generalizate' acumularea de mari cantitati de lic+id in pleura sau
peritoneu. Hipovolemiile sint insotite sau nu de modi#icarea H4. 3n +emoragiile acute' la
inceput'+ipovolemia este normocitemica' apoi' pe masura intravazarii de lic+. interstitial' Ht
incepe sa scada si +ipovolemia devine oligocitemica. 3n cazul pierderilor de plasma si/sau
lic+ide electrolitice' datorita +emoconcentratiei' +ipovolemia este policitemica. Daca se
pierd lic+ide +ipotone )transpiratii' varsaturi' diureze apoase*' plasma devine +ipertona'
+ematiile se ratatineaza' scade B2 si Ht scade' indicind ast#el pierderi mai mari ca in
realitate. 3n cazul pierderilor de lic+ide mai putin +ipotone' )bila' suc pancreatic* prin
varsaturi sau #istule digestive' sau in cazul evacuarilor repetate de transsudate pleurale
sau peritoneale' plasma devine usor +ipotona' +ematiile cresc in volum' creste B2 si
ast#el Ht indica pierderi mai mici ca in realitate. 3n +emoragiile acute datorate leziunior
venoase ce nu depasesc 10& din vol.sang+. total' presiunea arteriala )!5* practic nu se
modi#ica' iar in cazul leziunilor arteriale ce duc la pierderi de sub 10& din vol.sang+' !5
se modi#ica' dar pe termen scurt si in mica masura' datorita declansarii prompte a
mecanismelor compensatorii= mobilizarea singelui din depozite' intravazarea lic+.interst.
in capilare' oligurie pina la anurie' cu declansarea sistemului 0151aldosteron' scaderea
secretiilor e$ocrine' a,ustarea patului vascular la vol.sang+. scazut prin vasoconstrictie
selectiva si redistributia singelui spre organe vitaleG se sunteaza sectorul arteriolo1venular'
deci scade patul vascular.
Daca +emoragia se soldeaza cu pierderea de peste 80& din Bol sang+' se instaleaza
socul. Acesta se instalea)a in hipovolemii asolute, datorate pierderilor lichidiene si
in hipovolemii functionaleA decompensarea acuta a B( in cazul 3C5' tamponadei
pericardice' rupturii de valve cardiace' in toate aceste cazuri aparind /!5 )edemul
pulmonar acut*.
$*&.Cipervolemiile normocitemice,oli'ocitemice si policitemice
<ipervolemiile sau pletorele sang+ine se pot instala datorita cresterii B! in cazul
hipervolemiilor oligocitemice' cind B2 este - sau usor scazut asa cum se intimpla in
retentiile +idrosaline= la pacientii cu insu#icienta renala' la cei cu ciroza si la cei cu
insu#icienta cardiaca.
<ipervolemia policitemica se caracterizeaza prin cresterea B2' #ara modi#icarea B!=
poliglouliile secundare )altitudine*' boli respiratorii cronice sau poliglouliile
primareA Policitemia /era.
<ipervolemia normocitemica se datoreaza cresterii atita B! cit si a B2' cu Ht -=
+ipertiroidie' trans#uzie de singe integral. Hipervolemiile acute se datoreaza per#uziei
masive de di#erite solutii= singe' plasma' masa eritrocitara' solutii cristaloide.
#epartitia volumului sanghin este inegala, cea mai mare parte' %5& gasindu1se in
sistemul de ,oasa presiune' adica in re)ervorul de capacitate, reprezentat de cord' venele
mari' capilarele din marea circulatie' mica circulatie' iar restul de 15& sint repartizate in
aorta si artere' ce reprezinta re)ervorul de presiune.
3n conditii #iziologice si in repaos' ,umatate din B( total se a#la e#ectiv in circulatie' ca
B( circulant' iar restul circula de10112 ori mai lent' sau stagneaza in sectoare ale
sistemului venos' ca B( de rezerva. 3n #icat e$ista 0'511L de singe' in splina 0'810'5L' si de
asemenea' la nivelul ple$urilor subpapilare. La aceste nivele' se pot acumula cantitati
relativ mari de (' #ara modi#icarea presiunii intravasculare
?A
$*(.repartitia volumului san'uin
$*).controlul volemiei
"ontrolul volemiei. Cecanismele de baza ale controlului volemiei sint aceleasi care
controleaza !5. 3n cazul cresterii volemiei' creste debitul cardiac' creste !5' creste
presiunea in a.renala' creste #iltratul glomerular' creste diureza si ast#el se corecteaza
volemia.
-atriureza si diureza variaza direct proportional cu valoarea !5. !t valori mici ale
volemiei' mecanismul invers' de retentie +idrosalina are la baza sistemul 0155ldosteron.
3ntervin rapid )secunde' minute* re#le$ul baro1receptor arterial si cel de
intindere a receptorilor din zonele de ,oasa presiune )Estretc+ receptorsF*' denumit Ere#le$
de volumF' ast#el= cresterea volemiei duce la cresterea !5 in sistemul aortic si la cresterea
presiunii in 5.pulmonara si alte zone intratoracice de ,oasa presiuneG ast#el sint e$citati
baroreceptorii sino1carotidieni si aortici si receptorii de intindere din zonele de,oasa
presiune' ca urmare este in+ibat (-simpatic' si are loc vasodilatatia la nivelul arteriolelor
renale' ast#el crescind diureza.
Hipervolemia stimuleaza si eliberarea de atriopeptina, ca urmare va creste natriureza si
diureza de 8 pina la 10 ori' mecanismul ne#iind' insa' de lunga durata. 7oncomitent'
atriopeptina in+iba eliberarea de aldosteron.
3n cazul +ipovolemiei' aldosteronul #avorizeaza reabsorbtia de -a si apa in segmentele
distale ale tubilor renali' ca urmare volemia creste cu 10120& in primele doua zile dupa
cresterea eliberarii de aldosteron. !e masura ce creste !5' datorita cresterii volemiei' se
instaleaza #enomenul Ealdosteron escapeF care consta in cresterea diurezei si natriurezei
in ciuda aldosteronului eliberat in cant mai mare. Bolemia revine in citeva zile la valori
cu 5110& mai mari ca normal' dupa #enomenul Ealdosteron escapeF.
3n mentinerea volemiei intevin si eliberarea de 5DH si mecanismul setei' ca si
mecanismul Fapetitului pentru sareF' ce intervine in +ipovolemia din 6.5ddison' neuronii
din structurile ce regleaza apetitul pt.sare #iind situati in aria anteroventrala a B8. Lezarea
acestora )traumatica' isc+emica' e$perimentala* induce disparitia apetitului pentru sare.
Centinerea normala a volemiei se datoreaza si sc+imburilor lic+idiene de la nivelul
capatului arterial al capilarului' interstitiului si capatului venos al capilarului' guvernate
de un ,oc presional denumit EFortele (tarlingF. !resiunea +idrostatica la capatul arterial
este de 82mmHg' cea coloid1osmotica de 2% mmHg' presiunea coloid1osmotica din
interstitiu este de 5 mmHg' iar cea +idrostatica a lic+idului interstitial este negativa'
datorita suctiunii lim#atice. (tructurile solide din interstitiu dezvolta o presiune de sens
contrar celei din lic+id' iar turgorul cutanat )elasticitatea pielii* e$ercita o presiune de 2
mmHg. (uma algebrica a #ortelor de la capatul arterial al capilarului este egala cu o
presiune e#ectiva de #iltrare de citiva mmHg' al carei e#ect este e$travazarea de lic+id din
capilar in interstitiu= 0'8& din plasma capilara. La capatul venos' predomina #orta de
intravazare' datorita scaderii presiunii +idrostatice la 15mmHgG reintra in capilar 9/10 din
totalul lic+idului e$travazat' iar 1/10 ia calea lim#ei.
833.vascozitatea sanelui
/isco)itatea (/) singelui este un element al rezistentei intravasculare la curgerea
singelui' pt.ca singele are in conditii #iziologice un regim de curgere neturbulent.
/cuatia Hagen1 !oiseuille' N )#lu$ul* = delta ! $ 0 %//S TL. Biscozitatea depinde de
#actori plasmatici )globuline al#a 1' al#a 2' beta si gama*' #ibrinogen. 3n a#ibrinogenemii'
B scade #oarte mult' iar in +iper#ibrinogenemii' creste #.mult. Biscozitatea variaza direct
proportional cu Ht. 3n +iperviscozitate' scade mult viteza de circulatie a ( in
microcirculatia +epatica' cerebrala' +epatosplenica si e$ista tendinta la agregare
intraarteriolara a +ematiilor' ceea ce duce la isc+emie si staza in acele teritorii' urmate de
semne neurologice de #ocar ce sugereaza un accident vascular cerebral isc+emic si semne
?C
de retinopatie. Hiperviscozitatea sanguina are consecinte #oarte grave= in#arcte #ara
tromboza' gangrena localizata #ara ocluzie arteriala. HiperB prin cresterea Ht este
intilnita in !olicitemia Bera. HiperB este provocata si de cresterea agregarii eritrocitare
in di#erite situatii' precum sint= arsuri' into$icatii' embolii grasoase' trans#uzii masive de
singeG +iperB prin scaderea temperaturii ambiante este intilnita in +ipotermii spontane
sau provocateG HiperB prin scaderea de#ormabilitatii eritrocitare se intilneste in di#erite
+emoglobinopatii si alterari ale metabolismului eritrocitar. Hipoviscozitatea se intilneste
in anemii' +emodilutii' +ipoproteinemii' +ipertermii.
83L.(unctiile sanelui
Functiile singelui. Functia respiratorie, datorita transportului gazelor respiratorii'
Functia nutritiva, datorata transportului de subst.energogenetice'plastice de la nivelul
tubului digestiv la nivel tisular' si de la organele de depozit la alte tesuturi.
Functia e8cretoare datorata transportului catabolitilor la organele de eliminare )plamin'
rinic+i' piele' intestin' #icat' gl.sudoripare*.
Functia de control al ech.hidroelectrolitic, datorata sc+imburilor dintre plasma'
lic+.interstitial si celule' care asigura izoionia )ec+ilibrul ionic*' izotonia )mentinerea
!.sm a plasmei* si izo+idria )ment.ec+.acido1bazic*. !t.realizarea acestei #unctii' singele
asigura transportul a numerosi +ormoni implicati.
Functia de termoreglare asigurata prin transportul de caldura de la organe cu
metabolism intens )F* la supra#ata corpului' ast#el realizindu1se termoliza. . importanta
aparte in aceasta #unctie o detine caldura speci#ica mare a (' datorita apei ce reprezinta
90& din compozitia singelui.
Functia de hemosta)a, realizata prin trombocite si o serie de #actori plasmatici
indispensabili +emostazei si #ibrinolizei'
Functia de aparare impotriva diversilor agenti patogeni si cea de eliminare a
elementelor celulare imbatrinite' realizata prin cooperarea unor mecanisme comple$e
imune' speci#ice si nespeci#ice' ce implica elemente #igurate ale singelui' elemente
tisulare si proteine diverse' cu origine plasmatice sau tisulara.
Functia de coordonare si reglare, alaturi de SB", asigurata de transportul +ormonilor'
mediatorilor c+imici' substante biologic active' toate implicate in mentinerea
+omeostaziei.
8LH.,ematopoeza em0rio(etala
<ematopoe)a (<) este de#inita drept proliferarea cel.progenitoare )precursoare*' al
caror numar e mentinut ct. de celulele stem si diferentierea lor in toate componentele
celulare ale singelui. Sediul < depinde de virsta individului si de starea #iziologica sau
patologica a acestuia. 3.:tapa emriofetala, alcatuita din > suetapeA -.:tapa
me)olastica, in care
eritropoieza este e$traembrionara si intravascularaG ast#el' in insulele sang+ine
mezodermale ale sacului vitelin' se observa in sapt.8112 de sarcina hemangiolasti,
cel.mezenc+imale sosite prin migrare din Elinia primitivaF a blastomerului
primitiv.3ncepind cu a 221zi' insule sang+ine asemanatoare' dar mai putin numeroase se
observa si in tes.mezoblastic al embrionului. 3nsulele e$traembrionare si intraembrionare
vor #uziona' alcatuind un sistem vascular unic. 3n aceasta etapa' se sintetizeaza Hb 2oUer
3'cu 2 lanturi zeta si 2 lanturi epsilon' care se va trans#orma rapid in Hb !ortland' cu
#ormula lanturilor globinice zeta 2 gamma 2 si apoi Hb 2oUer 33'cu 2 lanturi al#a si doua
lanturi epsilon. La #at se sint. Hb F' cu 2 lanturi al#a si 2 lanturi gamma.
!.:tapa hepatosplenica se caracteri)ea)a prin migrarea celulelor pluripotente la
acest nivel, unde e$ista conditii superioare pt.+ematopoeza. /a ocupa intervalul cuprins
intre sapt.91luna 5' spre s#irsitul caruia +ematopoeza scade la acest nivel'dar se mentine in
grad redus pina la 2 sapt.dupa nastere. La nivel splenic' +ematopoeza se prelungeste pina
??
in luna % de sarcina.
>.:tapa medulara incepe sa fie activa din luna = a /&3, iar incepind cu luna 7' devine
principalul sediu al H.
8L8.eritropoeza la adult
La adult, sediul < se limitea)a la nivelul
oaselor scheletului a8ial -vertebre'coaste' stern' bazin' oase craniene' epi#ize pro$imale
#emur si +umerus. Caduva rosie este activa' #unctionala' iar cea galbena apare prin
inlocuirea celulelor specializate cu adipocite. La 20 de ani' inlocuirea este completa'
realizindu1se centripet' dintre e$tremitati spre trunc+i. 5dipocitele medulare di#era mult
de cele din tes.adipos pr.zis= ele sint cel.adventiceale' reticulare si macro#age
trans#ormate prin incarcarea cu grasime' si detin si alte roluri' in a#ara celui de sustinere.
Populatia celulara a maduvei hematogene este alcatuita dinA celule stem
pluripotente, care sint cele mai timpurii. (int desemnate prin prescurtarea 7F<1(
)colonK #orming units1spleen*G aceste celule #ormeaza colonii pure si mi$te de
cel.precursoare la nivel splenic' la soareci iradiati. Cor#ologic' aceste celule sint asemanatoare
lim#ocitelor mici'dar sint incomplete din punct de vedere imunologic. La
om e$ista 7F<1blast' celule pluripotente' asemanataoare celulelor 7F<1(. Din aceste
celule primordiale' se dezvolta 2 linii celulare=
-."elule stem multipotente limfoide' din care deriva lim#ocitele 4 si 6 si
2."el stem multipotente mieloide' din care deriva precursorii pentru seria elementelor
rosii )+ematii*' granulocite' monocite si megaIariocite.
CH se caracterizeaza prin particularitati de microclimatG stroma celulara a CH contine
cel.endoteliale' #ibroblasti' si cel.adventiceale' macro#age centrale' denumite Enurse
cellsF' cel.doica ce asigura Fe necesar sintezei Hb de catre eritroblastisi' cel.adipoase
speci#iceG matricea e8tracelulara contine glicoproteine ce act.ca situsuri de recunoastere
si de adeziune pt.cel.stem +ematopoetice' precum= #ibronectina' laminina'
colagenul'proteoglicanii.
8L2.particularitatile meta0olismului eritrocitar
(etaolismul eritrocitar. /ritrocitele sint saci cu Hb' a caror principala #unctie
este transportul gazelor respiratorii' ce are o mare componenta pasiva= di#uziunea gazelor.
/ritrocitele sint lipsite de nucleu' mitocondrii si 0/' metabolismul lor #iind #oarte scazut=
ele participa cu doar 0'1& le metabolismul energetic general )2cal/2ore*. (ingura
sinteza realizata este a glutationului.
<ematiile necesita energie pentruA mentinerea formei, a deformailitatii
eritrocitare, asigurarea concentratiilor fi)iologice de o parte si alta a
m.eritrocitare, a electrolitilor (Ba, C), mentinerea Fe in stare feroasa, mentinerea
gruparilor sulfhidrice S< in stare activa, redusa, stare necesara pentru fi8area si
elierarea de $! si "$!. %oate aceste necesitati sint asigurate e8clusiv prin
metaoli)area gluco)ei. 100mL de masa eritrocitara consuma 0'810'mg glucoza/ora.
5lterarea procesului
de energogeneza eritrocitara duce la modi#icarea #ormei eritrocitului= s#erocit' ec+inocit'
stomatocit' sau la modi#icari ale #unctiilor acestuia= o$idarea Hb. 3n toate aceste situatii'
apare #enomenul de +emoliza patologica' intravasculara' adica anemia +emolitica de
cauza corpusculara.
8LA.tipuri de ,emolo0ina normala
<emogloina. Fiecare eritrocit contine !6->.pg<. (<:(). La o volemie de 5L de
singe' si o concentratie de 15gHb&' e$ista %00g Hb. 2C este de 9500. 3esita din
+ematie' Hb este #iltrata la nivel renal sau rapid catabolizata de celulele (0/. /ste o
+emoproteina tetramerica' alcatuita 90& din globina si 10& din gruparea prostetica'
+emul' o metalopor#irina ce contine Fe;;. La adult' Hb5 contine 2 lanturi al#a cu 11aa
?3
si doua lanturi beta cu 19aa' legate prin legaturi labile' punti de H sau legaturi Ban der
Qaals. 7ele lanturi realizeaza o structura globulara compacta' ce are in vecinatatea
supra#etei e$terne cavitati' pungile +emului. /$ista zone de contact intre lanturile
subunitatilor vecine' al#a11beta2 si al#a21beta1. 3n a$ul moleculei e$ista o cavitate
centrala cu 2 mici #osete' ce separa lanturile al#a si beta' #osete in care se #i$eaza 2'8D!2.
8LC.senescenta eritrocitara
Senescenta eritrocitara. Jilnic' 890 miliarde de +ematii sint indepartate din circulatie.si
inlocuite cu elemente tinere. Durata de viata a +ematiilor este de 1151120zile. !e masura
ce imbatrinesc' +ematiile su#era o serie de modi#icari= au tendinta la s#ericitate' scade
continutul lor in D;' 7a;;' esteri #os#orici' apa' lipide totale' colesterol' 54!' 2'81D!2'
grupari (H' scade utilizarea glucozei' scade activitatea enzimatica' scade de#ormabilitatea
eritrocitara' creste #ragilitatea mecanica si osmotica' creste continutul de metHb' ca
urmare a reducerii activitatii metHbreductazei si dia#orazelor' creste continutul de
Hb52'cu a#initate crescuta pt..2' creste viscozitatea interna eritrocitara' creste continutul
de -a;' scade #os#orilarea spectrinei' scade progresiv continutul de acid sialic' adevarat
EcenzorF al duratei de viata a +ematiilor.4oate glico#orinele din mb.eritrocitara su#era
desialinizare. Hematiile imbatrinite e$pun pe supra#ata mb. factorul de senescenta
eritrocitara, un glicopeptid pre)ent doar pe m.hematiilorimatrinite, ce nu contin
acid sialic. 5cesta este recunoscut de macro#agele splenice. Hematiile imbatrinite mai
e$pun pe versantul e$tern al mb. fosfatidilserina, ce repre)inta inca un semnal de
recunoastere de catre macrofage a hematiilor imatrinite. "aptarea hematiilor
imatrinite de catre macrofage se face printr-un sistem eta-galacto lectinic,
propriu acestora.
8L?.meta0olismul +erului
(etaolismul Fe. -ecesarul zilnic este de 20125mg. Hb contine cea mai mare parte de
Fe din organism' 70&' adica 2g. 0estul este stocat ca #eritina sau +emosiderina' o mica
parte este continut in mioglobina si o parte este legat de transportorul plasmatic'
trans#erina' alcatuind compartimentul labil. 5proape tot Fe este reciclat din catabolizarea
+ematiilor imbatrinite. Doar 5& din necesarul de Fe pt.+ematopoeza provine din tractul
gastrointestinal' absorbtia zilnica #iind de 1mg/zi. !ierderile zilnice de Fe sint realizate
prin #ecale' urina' transpitatie' #anere )descuamare tegumentara*.
5bsorbtia de Fe ma$imala are loc in duoden si ,e,unul pro$imal. pH1ul gastric
reduceFe;;;alimentar la Fe;;. 7oncentratia Fe in citosolul celulelor mucoasei
intestinale reprezinta EbarieraF pentru trecerea Fe in plasma' e#ectul avind o latenta de
citeva ore. De la nivelul celulelor din tractul digestiv' Fe este preluat de transferina, o
proteina cu ! situsuri delegare a Fe, cara se masoara drept 4367 )4otal 3ron 6inding
7apacitK*. .data eliberata de Fe' trans#erina este
transportata inapoi in plasma de catre macro#age. 4rans#erina este si o proteina de
transport citosolic (tocarea Fe se #ace in macro#age' ca feritina, un amestec +idrosolubil
de Fe;;' .H1 si .2' invelit de o proteina' apoferitina.
8L3.,emoliza +zioloica
<emoli)a fi)iologica )eritroliza* are ca principal splina' care impreuna cu #icatul' CH'
gg.lim#atici' monocitele circulante si macro#agele tisulare alcatuiesc (0/. (ediul
eritro#agocitozei este pulpa rosie splenica. /tapele acesteia sint= aderenta +ematiei la
supra#ata macro#agului' inglobarea si digestia prin #ormare de #agolizozmi' ce contin
+idrolaze lizozomale. 5cestea distrug in citeva minute eritrocitul inglobat. Hematiile
normale sint #agocitate #ara #i #ragmentate in preralabil' iar cele patologice sint initial
#ragmentate' apoi #agocitate. Din catabolizarea Hb rezulta globina' +idrolizata in aa ce
vor intra in #ondul comun metabolic de aa' proces responsabil de mari pierde energetice'
ce reprezinta o importanta parte din C6' si +emul' care este convertit in bilirubina' printrun
?L
comple$ proces enzimatic. /ritroliza #iziologica duce la eliberarea in plasma a unei
mici cantitati de Hb' care este legata de o al#a21glicoproteina de origine +epatica'
denumita haptogloina. 7omple$ul Hb1+aptoglobina este captat si metabolizat la nivelul
celulelor din (0/' mai ales la nivel +epatic.
8LL.relarea eritropoiezei
#eglarea eritropoe)ei. (e realizeaza predominent prin mecanisme de tip #eed1bacI
asupra celulelor nedi#erentiate' semnalele declansatoare provenind de la nivel tisular'
datorita variatiei aportului de o$igen. 5cest tip de #eed1bacI are drept element declansator
produsii de degradare ai Hb' aparuti in conditiile unei +emolize intense'
e#ectul #iind de stimulare a eritropoezei.
0eglarea umorala a eritropoezei este realizata de eritropoetina si de factorii de crestere
necesari regenerarii cel.stem (3L%&-"SFA &L>, o glicoproteina cu 2C20180Ddaltoni'
importanta pentru toate liniile celulare' ce actioneaza asupra cel.pluripotente timpurii'
pentru autointretinere )+ematopoeza de tip +omoplastic* si pentru di#erentierea
cel.pluripotente' in celule di#erentiateEcomiseF)+ematopoeza +eteroplastica*. 3L8 este
esentiala la nivelul CH pentru asigurarea celulelor responsive la eritropoetina. 3l8 este
sintetizata de L4' celule stem si are drept celule tinta celulele multi17(F.
8LD.eritropoietina
:ritropoetina (:) este produsa 90& in corticala renala si 10& in #icat' eliberarea sa in
plasma #iind in principal raspunsul la +ipo$ie al celulelor epiteliale glomerulare renale.
!recursorii timpurii ai eritrocitelor nu au receptori pt /' ci pt.3L8. !roeritroblastii )7F</1
colonK1#orming1units1erKtroid* sint primele celule din seria rosie cu receptori pt./. !rin
#eed1bacI negativ' cind scade presiunea partiala .2 in corticala renala' creste producerea
si eliberarea de / in circulatie.
!roducerea de / este stimulata de +.androgeni' alcaloza' catecolamine prin
mec.betaadrenergic.
: initiaza sinteza de Hb la nivel medular' scurteaza timpul de tranzit medular
al eritroblastilor in curs de maturare si stimuleaza eliberarea de reticulocite in singele
peri#eric. 5nemii prin nivel scazut de / e$ista in= 30cronica' +ipotiroidie )dat.scaderii
consumului de .2*' +ipopituitarism )prin scaderea de gonadotropi*' +ipoparatiroidism
)dat.calci#icarilor renale corticale*. #eglarea nervoasa a eritropoe)ei este realizata prin cale
re#le$a' prin stimularea
c+emoreceptorilor sinocarotidieni' e#erentele bulbare adrenergice determinind prin
intermediul n.span+nici la nivelul CH cresterea eliberarii de reticulocite si +ematii adulte
in singele peri#eric.
8LG.polilo0uliile
Poliglouliile. Policitemia vera se caracterizeaza prin cresterea nr.de +ematii'
trombocite si neutro#ile' ea putind evolua spre leucemie granulocitara cronica acuta si
deces in cea de a treia etapa de evolutie a bolii. (emnele clinice si paraclinice ma,ore sint
splenomegalia' a#ectarea severa a microcirculatiei )cerebrale'+epatosplenice*' cresterea
Ht si a nivelului de ac.uric in plasma.
Poliglouliile secundare apar in cazul= #umatorilor )prin cresterea compensatorie a
nivelului de /*' Hbpatiilor si +ipo$emiilor cr.'precum sint bolile respiratorii cr.' (d.de
apnee in somn;obezitate )(d. !icIUeeI*' boli congenitale cardiace cianogene cu sunt drstg.
5lte situatii sint cele in care e$ista e$ces de /= carcinom renal' rinic+i polic+istic'
+idrone#roza' re,etul de gre#a renala )mecanism inca neelucidat*' tumori e$trarenale
secretante de / situate la nivel +epatic )sindrom paraneoplazic*' #eocromocitom' tumori
cerebeloase' leiomiom uterin de mari dimensiuni )benign*'care prin compresie mecanica
asupra a.renale' duce la e$ces de /. 3n (d.7us+ing si in cazul aportului e$ogen de
androgeni' creste eliberarea de /.
?D
8DH.antienele sistemului A>E
Antigenele sistemului A$D. 3n /uropa' ordinea descrescatoare a prezentei ag.5 si 6.pe
supra#ata +ematiilor #ace ca grupa de singe cea mai #recvent intilnita sa #ie 5' urmata de
grupele . si 6' pentru ca cel mai mic procent de indivizi sa apartina grupei 56.
5g.sistemului 5.6 se a#la pe supra#ata tuturor celulelor din organism' precum si in
di#eritele lic+ide biologice. /le apar din luna % a vietii 3< si devin complet mature la 9112
luni dupa nastere. 6ioc+imic' aceste ag.sint glicoproteine cu 2C 80000011
milion'alcatuite dintr1un sc+elet peptidic pe care sint #i$ate covalent numeroase lanturi
oligoza+aridice. 7aracterele ag1ice ramin nemodi#icate daca ag1ele sint tratate cu enzime
proteolitice )papaina*. Daca sint tratate cu enzime glicolitice de origine bacteriana sau
parazitara )protozoare*' dispare speci#icitatea ag1ica initiala ceea ce demonstreaza ca
raspunzatoare pentru aceasta sint radicalii glucidici. Din sistemul 5.6 #ac parte
antigenele 5' 6 si H. /le deriva dintr1un precursor comun' cu structura lipopreoteica' ce
are atasat un lant de monoza+aride' ultimul #iind galacto)a. 5st#el' in cadrul sistemului
5.6 e$ista 9
genotipuri )55' 66.56' 5.'6.'..*. 3ndivizii apartinind grupei . sint lipsiti de
antigenele 5 si 6' dar au pe +ematii ag.H. Fenotipic' e$ista doar variante' deoarece
55=5. si 66=6.. Locusul genetic 56. contine si alte doua gene' 51 si 52.
8D8.alutinele sistemului A>E
Aglutininele sistemului A$D sint anticorpii naturali si anticorpii imuni anti A si anti
D, denumiti aglutininele alfa si eta. "ei naturali (aglutinine) apar in plasma in
absenta aglutinogenului )ag* eritrocitar' titrul la nastere #iind 0. (inteza lor incepe sa
apara intre lunile 21% dupa nastere' titrul creste lent pina la virsta de %110 ani' raminind
apoi nemodi#icat pina la senescenta' cind scade lent. 5ceste aglutinine sint
gammaglobuline sintetizate de celule imunocompetente si apartin claselor 3gC si 3g2.
Anticorpii imuni apar in plasma dupa trans#uzii de singe incompatibil. /i au putere
aglutinanta mai mare si in prezenta complementului au actiune +emolitica' #iind denumiti
hemoli)ine.
8D2.antienele sistemului 6,
Antigenele sistemului #h sint independente de sistemul 5.6./le sint prezente pe
+ematii incepind cu luna 3 de viata 3<' genele ce codi#ica sinteza lor #iind pe
cromozomul1. 7el mai important ag.al acestui sistem este ag.D sau 0+' prezent pe
+ematiile a %5& dintre indivizi' denumiti 0+ pozitivi' restul de 15&' la care ag.D este
absent #iind 0+ negativi. 3 n a#ara acestuia' mai e$ista inca peste 80 de ag.apartinind
acestui sistem' dar #ara importanta practica. 5g.din acest sistem sint de natura
lipoproteica. 5bsenta tuturor ag.din acest sistem de pe supra#ata +ematiilor este denumita
E#enomen 0+ nulF si este asemanator #enotipului 6ombaK' in sensul restringerii
posibilitatilor de donare/primire de singe strict la membrii acestui #enotip. 5cesti indivizi
au o #orma particulara de anemie +emolitica' datorita scaderii duratei de viata a
+ematiilor' datorita modi#icarii proprietatilor membranei eritrocitare' ca e#ect al absentei
tuturor ag.din acest sistem.
8DA.anticorpi sistemului 6,
Anticorpii sistemului #h sint numai de tip imun' ei rezultind in urma izoimunizarii dupa
trans#uzii' sarcini incompatibile sau in urma autoimunizarii' la cei cu de#icit imun.7el
mai important ac.este ac.antiD, ce apartine 3g2 si traverseaza bariera #etoplacentara'
producind anemia hemolitica a nou-nascutului sau eritrolasto)a fetala,in cazul
#emeilor 0+ neg'a#late la a doua sarcina cu #at 0+ poz. 3n acest caz' al doilea contact al
+ematiilor #atului cu singele matern permite cresterea rapida a titrului de ac.antiD' in
cadrul raspunsului imun de tip secundar' anticorpi ce produc +emoliza la #at. !t a preveni
acest risc' se recurge la administrarea de anticorpi antiD' preparati din plasma indivizilor
?G
0+ neg' imunizati la ag.D. . doza= 1mL ce contine 800Vg' su#icienta pt.a contracara
e#ectul imunizant a 80mL de singe integral. (e administreaza pt. a preveni imunizarea
#emeilor 0+ neg.e$puse eritrocitelor 0+ poz.prin= +emoragii materno1#etale )avort'
metroragii in timpul sarcinii' sarcina ectopica' traumatism abdominal in timpul
sarcinii'amniocenteza*G in cazul primei sarcina cu #at 0+ poz' se adm.postpartum. La o a
doua sarcina' se adm.antepartum' la 2% sapt.de sarcina. Anticorpii sistemului #h sint numai
de tip imun' ei rezultind in urma izoimunizarii dupa
trans#uzii' sarcini incompatibile sau in urma autoimunizarii' la cei cu de#icit imun.7el
mai important ac.este ac.antiD, ce apartine 3g2 si traverseaza bariera #etoplacentara'
producind anemia hemolitica a nou-nascutului sau eritrolasto)a fetala,in cazul
#emeilor 0+ neg'a#late la a doua sarcina cu #at 0+ poz. 3n acest caz' al doilea contact al
+ematiilor #atului cu singele matern permite cresterea rapida a titrului de ac.antiD' in
cadrul raspunsului imun de tip secundar' anticorpi ce produc +emoliza la #at. !t a preveni
acest risc' se recurge la administrarea de anticorpi antiD' preparati din plasma indivizilor
0+ neg' imunizati la ag.D. . doza= 1mL ce contine 800Vg' su#icienta pt.a contracara
e#ectul imunizant a 80mL de singe integral. (e administreaza pt. a preveni imunizarea
#emeilor 0+ neg.e$puse eritrocitelor 0+ poz.prin= +emoragii materno1#etale )avort'
metroragii in timpul sarcinii' sarcina ectopica' traumatism abdominal in timpul
sarcinii'amniocenteza*G in cazul primei sarcina cu #at 0+ poz' se adm.postpartum. La o a
doua sarcina' se adm.antepartum' la 2% sapt.de sarcina.
8DC.eritro0las(aza (etala
Anticorpii de tipul /Y materni trec prin placenta si se leaa de ,ematiile (etale$
pe care le distru$ ceea ce determina anemia (etala$ ,iper0iliru0inemia !
cresterea cantitatii de 0iliru0ina in sanele (etal$ 0iliru0ina +ind produsul de
meta0olism al ,emolo0inei ! precum si cresterea celulelor eritrocitare tinere
compensatoare.
8D?.rolul (unctional al plac,etelor sanuine
8. /ntervin n men'inerea interit'ii endototeliului vascular$
2. #unc'ie de transport
A. #unc'ie de aprare
C. 6ol n in-ama'ie
?. 6ol n ,emostaz 1 #unc?ie esen?ial>' se realizaz> prin e$isten?a
#actorilor plac+etari )F*
F1 = accelerina )identic cu FB plasmatic*
W F2 = cu rol de accelerator al ac?iunii trombinei
W F8 = #os#olipide din membrana !' ec+ivalent
F333)lecitin>*
W F = anti+eparina
W F5 = serotonina
W F9 = #ibrinogenul plac+etar' ampli#ic> agregarea !
W F7 = trombostenina cu rol Xn retrac?ia
c+eagului'protein> contractil> din #amilia actinelor
F% = anti#ibrolizina plac+etar> cu rol de antiplasmin>
W F9 = #actorul stabilizator al al #ibrinei' consolideaz>
c+eagul de #ibrin>.
8D3.receptori trom0ocitari
8DL.timpul vascular
:ncepe odat cu lezarea vasului. "rima reac'ie const n vasoconstricia peretelui
acestuia$ produs at&t re-e% c&t i su0 ac'iunea serotoninei. Mrmeaz aderarea
trombocitelcr la nivelul plii$ aglutinarea i metamorfoza vscoas a acestora$
cu (ormarea unui trom0us al0 trom0ocitar care astup temporar vasul i duce la
oprirea s&nerarii n 2-C minute. Acest timp se prelunete n a(ec'iuni vasculare
sau trom0ocitare
3H
8DD.timpul plac,etar
Timpul plac,etar
4rombocitul' ''piticul mor#ologic' gigantul #iziologicF' nu ader> la
endoteliu -' intervine Xn +emostaza primar>
W aderare 1 adeziune
W activare
W agregare
W aglutinare
5D/050/5' se realizeaz> prin 0 speci#ici de pe ! 1 cls. integrine' ce
recunosc liganzi de pe alte celule' #b. de colagen denudate
Cai mul?i receptori de membran> se leag> de #actorii e$tracelulari ca
r>spuns la ruptura vasului de s@nge' rezult@nd aderarea Ai agregarea
plac+etelor. 5D!1ul Ai receptori membranari sunt ilustra?i pentru a ar>ta rela?ia dintre
stimularea 5D! Ai calea ac. 5ra+idonic cu eliberare de 4$52 care
stimuleaz> agregarea mai departe.
2enerarea trombinei Ai leg>tura supra#e?ei trombocitului cu #actorii cascadei
coagul>rii este de asemenea #igurate. 5743B50/5 " se datoreaz> interac?iunii ligant receptor' poate
#ii
1 intrinsec> )startul*
1 autoactivarea
8DG.(actori plasmatici ai coaularii
#A5T>6// "9ASMAT/5//
# / este +0rinoenul$ licoprotein de natur lo0ulinic ce se (ormeaz n +cat
# //$ protrom0ina se (ormeaz n +cat i este precursorul trom0inei. <ste o
licoproteic.
de natur lo0ulinic.
# /// trom0oplastina tisular sau trom0oSinaza este un comple% (os(o-lipido-
proteic ce se
sete n toate 'esuturile n special n creier i plm&ni.
# /V este reprezentat de 5a24el intervenind aproape n toate (azele coaulrii.
# V proaccelerina sau lo0ulina acceleratoare se (ormeaz n +cat$ este o
licoprotein.
<a se sete numai n plasma proaspt dispr&nd prin coaulare.
# V// proconvertina sau cotrom0oplastina se (ormeaz n +cat i este o
licoprotein de
natur lo0ulinic.
# V///este (actorul anti,emo+lic
A.<lconstituieuncomple%(ormatdin#V///c(componenta
coaulant) i (actorul von aille0rand (# va) o licoprotein sintetizat n
celulele endote-
liale i meacariocite +ind prezent n trom0ocite. 9ipsa # V///c produce o 0oal
transmis
ereditar denumit ,emo+lia A ce se caracterizeaz prin ,emoraii prelunite la
cele mai mici
traumatisme.
# /^$ (actorul anti,emo+lic E sau componentul trom0oplastinic al plasmei se
(ormeaz
n +cat i este o licoprotein de natur lo0ulinic. 9ipsa lui produce ,emo+lia E
0oal cu
simptome asemntoare ,emo+liei A.
# ^ sau Stuart-"ro_er se (ormeaz n +cat +ind de natur lo0ulinic. <ste un
(actor im-
portant n (ormarea comple%ului convertor al protrom0inei (protrom0inaza) care
intervine n
38
trans(ormarea protrom0inei n trom0in.
# ^/ antecedentul trom0oplastinic al plasmei sau (actorul anti,emo+lic 5este o
lo0ulin
care este leat de Sininoenul cu reutate molecular mare care are rol n
activarea # ^/.
# ^// (actorul de contact sau =aeman este de natur lo0ulinic i se activeaz
prin con-
tactul cu supra(e'e strine n prezen'a Salicreinei ini'iind mecanismul intrinsec al
coaulrii
dar i activarea sistemului +0rinolitic. "articip alturi de al'i (actori la activarea
Salicreinei care
va duce la (ormarea Sininelor din Sininoeni.
# ^/// sau (actorul sta0ilizator al +0rinei sau 9aSi-
9orandesteolo0ulinsintetizatn+cat
ce intervine n trans(ormarea +0rinei solu0ile n (orma sta0il insolu0il.
Ac'ioneaz i asupra
colaenului i +0ronectinei
8GH.calea intrinseca a coaularii
Mecanismul intrinseceste declanat prin expunerea plasmei la suprafee strine ncrcate
negativ. Aceasta va duce la activarea F XII prin intervenia kalicreinei. F XIIa n prezena kinino-
genuluicugreutatemolecularmareactioneazasupraFXIpecarelactiveaz.FXIaactiveaz
la rndu-i F IX.
FIXa mpreun cu F VIII i cu factorul 3 plachetar n prezena Ca formeaz complexul activator intrinsec al F X
5alea e%trinsec i intrinsec ce duce la (ormarea trom0usului sanuin.
6olul #^a i a trom0inei n procesul coaulrii
5A9<A /1T6/1S<5c
8. #ormarea "A5
2. #ormarea "i / al cii intrinseci
A. Activarea #^$ con-uien'a maBor i (ormarea "6>T6>ME/1A]</
8G8.calea e%trinseca a coaularii
Mecanismul extrinsec este iniiat de tromboplastina tisular (F III) provenit din esuturile
distruse ca urmare a leziunilor. Aceasta mpreun cu FVIIa prin intervenia F XIIa, IXa i Xa n
prezena Caformeaz un complex activator extrinsec al F X
5A9<A <^T6/1S<5c
8. Activarea #V//
2. #ormarea "i / al cii e%trinseci
A. 5on-uien'a maBor
5ctivarea ini?ial> a Fc 3Y de c>tre comple$ul 4F1B33a compenseaz>
de#icien?ele Xn #actori mai precoce ca FY33 Ai FY3
Zn+ibi?ia ulterioar> a 4F1B33a de c>tre 4F!3)calea in+ibitorie a #c tisular*
determin> ca activarea sus?inut> a FY de F3Ya Ai FB333a s> #ie critice pentru
+emostaza normal>' plac+ete Ai #os#olipid
8G2.rolul (c <"/ in cascada coaularii
8GA.(ormarea trom0inei
F Xa mpreun cu F V i cu fosfolipidele eliberate de trombocite i esuturi n prezena
Ca
2+
formeaz un complex cu aciune tromboplastinic sau protrombinazic ce va induce
transformarea protrombinei n trombin.
Trombina generat pe cale extrinsec, dei n cantiti mici, are rol n iniierea coagulrii.
Ea acioneaz i asupra trombocitelor favoriznd agregarea i activarea acestora.
Formarea trombinei pe cale intrinsec, ajutat de factorii plachetari, menine procesul
coagulrii prin transformarea unor mari cantiti de fibrinogen n fibrin.
Trombina care este o enzim proteolitic asemntoare tripsinei acioneaz asupra
fibrinogenului ajutnd la polimerizarea acestuia n fibre insolubile, proces favorizat de FXIIIa
n prezena Ca
2+
32
.
Formarea trombinei este un moment deosebit de important n procesul coagulrii nu
numai datorit aciunii directe asupra fibrinogenului, ci i datorit activrii factorilor V, VIII,XI
i XIII
"6>T6>ME/1A protrom0inaza dT6>ME/1c
indi(erent de oriine
P "rocesul are loc la supra(a'a plac,etelor n prezen'a 5a
5&nd este leat de trom0omodulin$ su(er o modi+care con(orma'ional care-i
crete a0ilitatea de a
activa proteina 5.
:n prezen'a proteinei S li0ere$ proteina 5 activat in,i0 coaularea ac'ion&nd
asupra #Va i V//a.
#r modi+care con(orma'ional trom0ina poate ac'iona asupra +0rinoenului !
dop de +0rin.
8GC.(ormarea +0rinei
Formarea filamentelor de fibrin organizate ntr-o reea n care sunt prinse i eritrocitele
ajuta la transformarea cheagului alb format din trombocite n cheag rou, mult mai rezistent,
care acioneaz ca un adevrat dop hemostatic.
Structurarea cheagului prin micorarea spaiului dintre filamente i concentrarea gelu-
lui vor duce n aprox 1 or la diminuarea sa n volum cam cu 15% constituind fenomenul de
sinerez. Urmeaz retracia cheagului prin care acesta i reduce volumul la doar 1/6 din cel
iniial prin expulzia serului devenind mult mai dens
Fibrinoliza are rol i n reparaia tisular, matricea rezultat n urma procesului ajutnd la
formarea esutului conjunctiv, proliferarea fibroblatilor i dezvoltarea vaselor sanguine.
Activatorii plasminogenului sunt prezeni n esuturi, snge sau urin.
Activatorul tisular este sintetizat de celulele endoteliale vasculare de unde este eliberat sub
influena unor stimuli (anoxie, staz, acidoz, catecolamine, vasopresin, trombina)
Activatorul urinar este activat sub aciunea plasminei sau a kalicreinei.
Exist i activatori exogeni cum este streptokinaza produs de streptococul hemolitic
folosit n practica medical pentru lizarea trombilor vasculari.
Inhibitorii fibrinolizei sunt reprezentai de substanele cu aciune inhibitoare asupra
Are loc En > etape:proteolitic$ polimerizarei sta0ilizare
W 1.(ub ac?iunea proteolitic> a trombinei' #ibrinogenul este scindat> Xn
#ibrinopeptide 5Ai 6 Ai monomeri de #ibrin>
W 2. !olimerizarea spontan> a monomerilor de #ibrin> =112s
W 8.4rans#ormarea #ibrinei solubile'dezintegrabil> Xn #ibrin> insolubil>
sub ac?iune #c = FY333' F8' 7a Ai trombin>
8G?.timpul trom0odinamic
Dup coaulare are loc$ su0 ac'iunea trom0osteninei plac,etare$ un proces de
retracie a cheagului. Din c,ea este e%pulzat un lic,id l0ui numit ser. Serul
este plasma (r +0rinoen i protrom0ina ce s-au consumat n procesul de
coaulare. 6etrac'ia c,eaului dureaz 2e2C ore. Dup retrac'ie$ c,eaul su(er
treptat un proces de topire numi brinoliz. Aceasta se datorete unei enzime
proteolitice$ plasmina (+0ri-nolizina)$ activat i ea n timpul coaulrii. #i0rinoliz
are drept e(ect ndeprtarea c,eaului i deso0struarea vasului prin care se
poate relua circula'ia. :n (elul acesta au (ost :ndeprtate toate consecin'ele lezrii
vasului
Tul0urrile ,emostazei pot aprea n oricare din cei trei timpi. Aceste tul0urri se
numesc sindroame ,emoraipare.
Activarea +0rinolizei se produce simultan cu activarea coaulrii$ prin (actori
comuni. Ast(el$ (actorii ^// i ^/ activa'i$ stimuleaz i activatorii +0rinolizei
"rocesul de coaulare evolueaz mai rapid$ iar cel de +0rinoliz mai lent
Dou> procese = 8) sinerez i 2)retrac'ia c,eaului' n ambele
procese se elibereaz ser
8G3.controlul coaularii-enumerati (actorii$ rol
P #lu%ul san$ contri0uie prin e( sale mecanice la (ram. +0rinei care
3A
ulterior este distrus.
P S0st. in,i0itorii naturale :
! <"/ (T#"/)$ in,i0itorul cii mediate tisular i a "i/ e%trinsec
! antitrom0inele plasmatice$ sunt mai multe dar cunoscute sunt
AT : /$ //$ /// i /V
8GL.antitrom0inele ! rol$enumerare
Antitrom0ina /// ! learea ei de ,eparin i mrete capacitatea
de a se lea i de a inactiva serinproteazele
AT ///
W are e(ect de protec'ie a ,eparinei (#Cpl)
W rol antitrom0oplastinic : (-) #/^a$V///a$ ^a
W (c' ca anti+0rinolizin
Activarea antitrom0inei ///
Heparina se #i$eaz> de lizina din molecula de 54 333 Ai induce modi#ic>rii
con#orma?ionale ale moleculei de 54 333
0estul argininic din molecula de 54 333 devine mult mai accesibil pentru
zona serinic> a trombinei.
!rin interven?ia +eparinei' reac?ia 54 333 cu trombina se accelereaz> de 1000
ori.
8GD.sistemul anticoaulant al proteinei 5
P /nclude :
P 2 proteine plasmatice : 5 i S$ se sintetiz.n +cat$ dep. de
vit. U
P i o protein din mem0rana cel endot vasc -
trom0omodulin
P nu se consum n coaulare
Activarea proteinei C la suprafaa endoteliului vascular.
Trombomodulina formeaz cu trombina un complex care n prezena Ca
scindeaz proteina C.
Proteina C este fixat la rndul su prin poriunea sa !amacarboxil!lutamic
(C"#) $i a Ca de o structur a membranei endoteliale
8GG.e%plorarea coaularii lo0ale si a etapelor
<%ist 2 ci de activare independente$ sistemul de contact Ai sistemul
mediat de #actorul tisular sau e$trinsic.
5cestea se Xnt@lnesc Xn momentul activ>rii FY Ai duc la generarea de
trombin> care trans#orm> #ibrinogenul Xn #ibrin>
5ceste reac?ii sunt reglate de antitrombin>' care #ormeaz> comple$e cu toate
proteazele serice ale coagul>rii cu e$cep?ia FB33 Ai de sistemul proteina 71(
care inactiveaz> #actorii B Ai B333
<^"9>6A6<A 5>AYM9c6//
%&%T'()# &*T+&*%'C 1 se determin> Xn ansamblu prin determinarea
47 sau 4 de recalci#iere a plasmei )4H*' !44)t. ce#alin>*' 5!44'
%&%T'()# ',T+&*%'C 1 se utilizeaz> 4N
-precierea transf .& / fibrin = prin 4 de trombin>
'xpl fazei de precoa!ul. 1 4 de consum al protromb.
La +emo#ilici acet timp se modi#ic> Xntruc@t #ormarea protrombinei
este de#ectuas> ' Xn ser r>m@n cant mari de F3' Xn sc+imb 4( este
normal
4<L6<0[03 D/ 7.52<L50/
W 0ipocoa!ulabilitate = +emo#ilia 5 Ai 6' dob@ndite = de#icit de vit D'
dis#unc?i +epatice' auto5c
W 0ipercoa!ulabilitate = tromboze asociat cu embolie
2HH. +0rinoliza ! de+nitie$ rol
F&D#&B$L&@A
P "c independent de ,emostaz$ dei ac'iunea sa se des(oar n st&ns
3C
corela'ie
P #i0rinoliza eed des(acerea enzimatic a +0rinei n (ramente ce nu
mai sunt capa0ile s men'in o re'ea
P Enzima implicat este plasmina
P "rocesul ncepe din centrul c,eaului$ iar la peri(erie sunt in,i0itori
+0rinolizei
P Din acest proces rezult repermeabilizarea vasului s!
P #ol - men'ine permea0ilitatea vaselor$ tu0ilor renali
- cur' (ocarele postin-amatorii
- controleaz concentra'ia plasmatic a (actorilor coaulrii :
distrue +0rina$ #V$ #V///$ protrom0ina c&nd este n cantitate crescut.
:n cantit'i mici releaz concenta'ia lor ca i (rac'iunii ale
complmentului$ A5T=
2H8.activatorul tisular al plasminoenului t"A
Activatorul tisular al plasminoenului )t!5* este eliberat din celulele
endoteliale' intr> Xn c+eagul de #ibrin> Ai activeaz> plasminogenul la
plasmin>
.rice cantitate de plasmin> liber> este comple$at> de in+ibitorul al#a 2 al
plasminei.
Fibrina este degradat> la #ragmente cu mas> molecular> mic>' produAi de
degradare ai #ibrinei )FD!s*
2H2.relarea +0rinolizei
Dup deradarea +0rinei$ plasmina rmas n circula'ie se com0in
rapid cu in,i0itorii naturali :
P -. &nhiitorii activatorului plasminogenului
! PAI, in,i0 t"A$ MU.
! in,i0itori sintetici : :A", A("<A
P 2. &nhiitorii plasminei
! 2 antiplasmina,
! 2 macroglobulina
! 2 antitripsina, antitripsina, ! " #antifibrinolizina$
2HA.imunitatea:de+nitie$clasi+care
8. condi'ie n care oranismul$ n contact cu un aent patoen (micro0ian sau de
alt natur) nu contracteaz n mod speci+c starea patoloic (0oala) indus de
aentul patoen respectiv.
2.totalitatea mecanismelor de aparare impotriva microoranismelor invazive
(0acterii$ virusuri$paraziti) si a structurilor non-sel(
/MM1/TAT<A /11AS5MTA 1<S"<5/#/5A: structuri si mecanisme cu rol protector
(teumente$ mucoase$ enzime din secretii e%ocrine ! lizozimul$ 0actericidina)$ p=-
ul secretiei astrice$ temperatura corpului ce o(era conditii precare de dezvoltare
unor microoranisme$ presiunea mare a >2 in alveolele pulmonare ce in,i0a
dezvoltarea ermenilor anaero0i$ nivelul cortizolemiei ce in-uenteaza raspunsul
in-amator si rezistenta la in(ectii
/MM1/TAT<A /11AS5MTA S"<5/#/5A: rezistenta prezenta la nastere unui oranism
(ata de anumiti aenti micro0ieni care il particularizeaza de alte oranisme din
aceeasi populatie sau specie$ sensi0ile la actiunea acelor aenti micro0ieni.
Atunci cnd rezisten'a caracterizeaz to'i mem0rii unei specii$ aceasta nu este o
condi'ie de imunitate$ ci o lipsa de suscepti0ilitate a speciei$ sau lips de
aresivitate a aentului micro0ian
/MM1/TAT<A D>E/1D/TA: condi'ia oranismului care prezint o stare de rezisten'
speci+c (a' de un anumit aent patoen micro0ian. Aceast rezisten' se poate
instala A5T/V$ ca /MM1/TAT< 1ATM6A9A$ n urma contactului cu di(erite
microoranisme$ sau A6T/#/5/A9A$ n cazul administrrii de antiene su0 (orma
VA55/1M6/9>6.
3?
/MM1/TAT<A 1ATM6A9A D>E/1D/TA /1 M>D "AS/V: este consecutiv pasaBului
anticorpilor materni la (t.
/MM1/TAT<A D>E/1D/TA A6T/#/5/A9: se poate instala n mod pasiv$ n urma
administrrii de seruri imune sau anticorpi pre(orma'i.
%.totalitatea mecanismelor de aparare impotriva
microor'anismelor invazive A-acterii, virusuri,parazitiB si
a structurilor non7sel
2HC.antienul-de+nitie$clasi+are$caracteristici
A1T/Y<1M9: este orice molecula de oriine endoen sau e%oen$ recunoscuta
de un oranism ca non-sel($ impotriva careia se declanseaza 6AS"M1S /MM1
speci+c umoral (productie de anticorpi) sau celular (clone de celule speci+ce
e(ectorii). De+nirea unei molecule de antien implica si capacitatea sa de a
interactiona speci+c cu e(ectorii raspunsului imun: anticorpi sau celule e(ectorii.
Moleculele cel mai 0ine recunoscute ca A sint proteinele cu masa moleculara
relativ mare (Mr Z 8HHHHD)$ dar pot (unctiona ca A si polipeptide de mici
dimensiuni si c,iar molecule de sinteza cu Mrf 8HHHD. "ot + A si moleculele
poliza,aridice$ lipidice$ acizi nucleici sau moleculele oranice de sinteza. 5ind in
mod accidental sint recunoscute ca A si moleculele normale ale propriului
oranism$ se instaleaza autoimunitatea
Antienul: caracter imunoen. imunoenitatea este complementar unei alte
calit'i a antienului$ speci+citatea: capacitatea antienului de a reac'iona
speci+c numai cu receptorii antienici solu0ili (anticorpi) sau cu receptorii
mem0ranari$ apru'i ca urmare a stimulrii antienice. /munoenele: antiene
caracterizate de imunoenitate i speci+citate. <le snt timodependente i
timoindependente.
A1T/Y<1<9< 5>M"9<T< T/M>D<"<1D<1T< declaneaz +e rspuns imun de tip
celular$ (6/5)$ +e rspuns imun de tip umoral (6/M). apari'ia rspunsului imun
este condi'ionat de implicarea 9/M#>5/T<9>6 T =<9"<6 (9T=) ntr-un proces
comple% de cooperare intercelular. MaBoritatea antienelor apar'in acestei
cateorii$ +ind de natur proteic
Determinan'ii antienici imunorelatori:
snt implica'i n controlul intensit'ii rspunsului imun declanat.
snt recunoscu'i de lim(ocitele T ,elper care ampli+ca rspunsul imun declanat
(determinan'i antienici =)$ sau de lim(ocitele T supresoare (determinan'i
antienici S)$ care limiteaz sau c,iar sisteaz raspunsul imun declanat.
Antienele snt caracterizate de imunoenitate$ speci+citate i de
imunomodulare! proprietate prin care antienul este implicat in decizia asupra
tipului de raspuns imun indus i a intensit'ii acestuia
2H?.oranizarea sistemului imun
Sistemul imun cuprinde douG compartimente2
$.compartimentul central2 alcGtuit din mGduva
+emato'enG Aunde se produc Ii matureazG limocitele 6
Ii se produc limocitele TB Ii din timus, sediul maturGrii
limocitelor T Ii
33
%.compartimentul perieric2 reprezentat de Fesutul
limoid capsulat Ii necapsulat. Splina Ii 'an'lionii
limatici alcGtuiesc Fesutul limoid capsulat Ii reprezintG
sediul derulGrii celei mai mari pGrFi a rGspunsului imun.
Ariile timodependente ale or'anelor limoide secundare
capsulate, populate de limocite T, sint2 paracorte1ul
'an'lionar si stratul limoid periarteriolar intern al
splinei. Ariile -ursodependente, populate de limocite 6,
ale acestor structuri sint2 corte1ul 'an'lionar si stratul
periarteriolar e1tern al splinei. 0elulele 6 sint dispuse in
a'lomerari denumite oliculi. Jesutul limoid necapsulat
este localizat la nivelul epiteliului di'estiv, -ronIic Ii la
nivel cutanat
2H3.celulele prezentatoare de antien
2HL.lim(ocitele T: clasi+care$activare
9/M#>5/T<9< T reprezint o clas celular (oarte ,eteroen$n cadrul creia se
descriu patru su0clase de lim(ocite T: helper! citoto"ice! supresoare i
contrasupresoare. Aceast clasi+care nu corespunde interal realit'ii$ ea
0azndu-se pe di(eren'ierea (enotipic a su0claselor lim(ocitare. Spre e%emplu$ nu
toate lim(ocitele T 5DC snt ,elper$ unele +ind citoto%ice sau c,iar supresoare$
aa cum nici toate celulele 5DD nu snt e%clusiv citoto%ice sau supresoare$ unele
avnd comportament de tip ,elper$ dovedit prin (actorii solu0ili secreta'i.
9im(ocitele T: reprezint 3H-DHR din totalul lim(ocitelor peri(erice i au o durat
de via' mare (ani$ zeci de ani)$ ele +ind intens recirculate. "e supra(a'a lor se
distin trei cateorii de receptori:
8.receptori pentru recunoaterea antienic$ i anume: T56 (T cell receptor).
comple%ul 5DA (5D K clas de di(eren'iere) cu rol n recunoaterea antienului
sau epitopului prezentat de 5"A. 5DC sau 5DD care snt co-receptoti implica'i n
recunoaterea moleculelor prezentatoare de antien M=5.
2.receptori cu rol accesor n activarea lim(ocitelor T: 5D2D$ 5DC?$ 5D2?.
A.receptori cu rol n adeziunea intercelular: 5D2$ 5D88@5D8D
9im(ocitele T recunosc antienul dup structura primar a determinantului
antienic$ spre deose0ire de lim(ocitele E i imunolo0uline$ care recunosc
antienele dup structura lor spa'ial. <le recunosc numai antienele e%puse
asociat de moleculele M=5 pe supra(a'a 5"A
Activarea lim(ocitelor T: este realizat numai de antienele T dependente$ n
cadrul unui proces ce necesit n mod o0liatoriu participarea 5"A. 5ooperarea
dintre 5"A i lim(ocit este de tip 0idirec'ional$ am0ele celule activndu-se reciproc.
Activarea lim(ocitului T de ctre 5"A este anterograd! ea derulndu-se n sensul
prezentrii antienului. Activarea lim(ocitului este realizat prin intermediul unor
semnale activatoare transmise spre citoplasm. Drept urmare$ au loc activarea
meta0olica$ e%pansiunea clonala precum si producerea de /9 (9T=) sau de
mediatori implicate in distruerea imunoenilor (9T5).
2HD.lim(ocitele E: distri0utie$rol in raspuns imun
9/M#>5/T<9< E reprezint 8?-2HR din totalul lim(ocitelor peri(erice. <le snt pu'in
recirculate i au durat scurt de via'. MaBoritatea snt distri0uite n corte%ul
anlionilor lim(atici i n pulpa al0 a splinei$ unde alctuiesc (oliculii lim(oizi. "e
supra(a'a mem0ranei lim(ocitelor E e%ist aceleai cateorii de receptori$ ca i n
cazul lim(ocitelor T: receptori pentru recunoaterea antienului E56 (E cell
3L
receptor)$ receptori cu rol accesor n activarea meta0olic a lim(ocitelor E$ (5DC?
i 5D2?) i receptori de adeziune intercelular. 9im(ocitele E
recunosc antienele solu0ile dup structura spa'ial a determinan'ilor antienici.
nu necesit prezen'a i implicarea 5"A n declanarea rspunsului imun.
dup stimularea antienic$ lim(ocitele E parcur etapele ciclului celular n mod
asemantor cu lim(ocitele T$ devenind lim(o0lati E$ a cror principal
caracteristic este prezen'a unui mare numr de molecule M=5// pe supra(a'a
mem0ranei celulare$ (apt ce le con(er posi0ilitatea de a (unc'iona ca 5"A.
celulele rezultate n urma diviziunilor celulare devin lim(ocite E e(ective$ adic
plasmocite$ cu direct implicare n derularea i +nalizarea rspunsului imun prin
sinteza de imunolo0uline. o mic parte dintre ele trec n rezerv ca lim(ocite E
cu memorie$ n anumite sectoare din oranele lim(oide secundare
9/M#>5/T<9< E pot Buca rolul de 5"A$ datorit prezen'ei pe mem0rana lor a
receptorilor de captare a antienelor (56$ #cg6 si E56) i a moleculelor M=5//$
prin care pot prezenta celulelor T diversele antiene. 5aracteristic lim(ocitelor E
este capacitatea lor de a capta antienele solu0ile i nu pe cele de tip
corpuscular.
2HG.6aspuns imun umoral
6AS"M1SM9 /MM1 MM>6A9 (6/M)
este declanat de antienele T-dependente.
necesit cooperarea a trei cateorii celulare: 5"A$ lim(ocitele T ,elper i T
supresoare$ (cu rol imunoreulator) i lim(ocitele E e(ectorii.
parcure dou etape$ prima cu e(ect activator$ derulat dinspre 5"A spre
lim(ocitele E$ care activate devin plasmocite productoare de anticorpi i a doua
etap cu e(ect in,i0itor sau de control$ care limiteaz rspunsul imun e+cient la
minimul necesar.cea de a doua etap este realizat prin intermediul unor
su0clase de imunolo0uline i prin activarea lim(ocitelor T supresoare care
limiteaz participarea lim(ocitelor E i T la 6/M. Men'inerea unui ec,ili0ru sta0il
ntre cele dou etape ale 6/M reprezint c,eia ,omeostaziei 6/M.
Secven'ial$ derularea 6/M parcure urmtorul model:
antienul declanator este preluat de 5"A (macro(ae sau celule dendritice).
transportat n ariile timodependente ale oranelor lim(oide$ este prezentat
lim(ocitelor T ,elper (T=)$ care se activeaz i devin lim(o0lati T=.
acetia mireaz n alte domenii ale oranelor lim(oide$ unde urmeaz s
ntlneasc lim(ocitele E cu care coopereaz.
simultan antienul este transportat pe cale lim(atic n zonele 0ursodependente
ale oranelor lim(oide$ unde este recunoscut de lim(ocitele E antien speci+ce$
care vor + activate$ tran(ormndu-se n lim(o0lati E ce mireaz treptat$ venind
n contact cu lim(o0latii T=$ cu care coopereaz (direct$ datorit numeroaselor
molecule M=5// de pe supra(a'a lim(o0latilor E sau indirect$ prin intermediul /92
i /9C$ eli0erate de lim(o0lastul T= activate).
in urma acestei cooperri$ lim(o0lastul E este complet activat$ el devenind
plasmocit productor de anticorpi speci+ci eli0era'i n circula'ie
6AS"M1SM9 /MM1: implic n mod a0solut necesar stimularea i proli(erarea
lim(ocitelor antien-speci+ce i sinteza unor molecule de recunoatere a
antienului$ reprezentate de anticorpi i@sau receptori mem0ranari.
> su0stan' este antienic dac ea declaneaz un rspuns imun i
reac'ioneaz speci+c cu anticorpii sau receptorii mem0ranari apru'i n timpul
rspunsului imun
6spunsul imun se deruleaz n trei secven'e:
8.#elecia clonal este etapa n care snt selectate de ctre antien doar acele
lim(ocite capa0ile s recunoasca antienul$ denumite lim(ocite antien-speci+ce.
3D
> clon lim(ocitar este o popula'ie de lim(ocite capa0il s recunoasca un
sinur tip de antien$ datorit prezen'ei pe supra(a'a mem0ranei celulare a unui
sinur tip de receptor pentru antien.
2.$ctivarea clonal% activarea meta0olismului intermediar al lim(ocitelor selectate
A.&"pansiunea clonal: proli(erarea celulelor selectate i activate meta0olic.
consecin': creterea numrului lim(ocitelor din clona stimulat de antien
28H.imunolo0ulinele-clasi+care$caracteristici
snt licoproteine identi+cate n plasm$ lic,idele intersti'iale i secre'iile
oranismului$ care au propriet'i de anticorpi$ ele com0inndu-se speci+c cu
antienele inductoare de rspuns imun.
structural$ imunolo0ulinele (/) snt alctuite din dou perec,i de lan'uri$ unele
cu reutate molecular mare Am0ele tipuri de lan'uri au secven'e varia0ile$
notate cu V= i V9 i pr'i constante$ notate cu 5= i 59. Secven'ele varia0ile V=
i V9 (ormeaz mpreuna situsul de recunoatere a antienului$ cu care acesta
sta0ilete leturi$ denumit paratop. > molecul de / con'ine dou situsuri de
leare a antienului identice$ capa0ile s recunoasc o unic structur
antienic$ prin urmare anticorpii snt structuri monospeci+ce.
Secven'ele constante ale / snt responsa0ile de celelalte (unc'ii ale acestora:
activarea complementului pe calea clasic.
ataarea la supra(a'a unor celule.
traversarea 0arierei (eto-placentare
Molecula de / are o varia0ilitate izotipic$ allotipica i idiotipic.
varia0ilitatea izotipic di(erentiaza mem0rii apartinind unor specii di(erite.
cea allotipic se datoreaz di(eren'elor enetice intraspecie.
varia0ilitatea idiotipic se datoreaz prezen'ei anumitor secven'e de aminoacizi
din reiunea varia0il a moleculei de /$ ce determin capacitatea de leare a
antienului. Varia0ilitatea idiotipic se coreleaz cu cea a situsului de leare a
antienului.
]ona varia0ila din structura /$ (denumita si idiotip) implicata in learea
antienului (#a0) poate (unctiona concomitent si drept determinant antienic.
Acesti determinanti antienici imunolo0ulinici$ aparuti in cantitate mare in
timpul 6/M determina la rindul lor aparitia de / anti-idiotip. Acest proces se
deruleaza in A-? valuri succesive$ ast(el realizindu-se o retea idiotip-antiidiotip$
implicata in stimularea sau in,i0area 6/M
/munolo0ulinele se mpart n ? clase (izotipuri)$ n (unc'ie de particularit'ile
secven'elor constante ale lan'urilor rele (5=)$ care snt de tip g$ b$Q$h$ i i.
/Y se resesc n concentra'iile cele mai mari$ ele reprezentnd L?R din totalul
/ circulante.
<%ist mai multe su0clase de /Y$ n (unc'ie de numrul de pun'i disul(urice
dintre lan'urile =: /Y8$ /Y2$ /YA i /YC. #unc'ional$ molecula de /Y con'ine
dou zone distincte: por'iunea #a0$ care este situsul com0inativ al moleculei de
/Y i este implicat strict n recunoaterea antienic i por'iunea #c$ denumit
(rament constant sau cristaliza0il$ responsa0il de ac'iunile 0ioloice ale
moleculei. /Y8 reprezint su0clasa dominant a /Y. Au rol important n
activarea complementului pe calea clasic$ n realizarea imunit'ii pasive a nou-
nscutului.
/Y2 este un sla0 activator al complementului
/YA este cel mai puternic activator al complementului pe calea clasic$ iar /YC
nu este activator de complement. /Y snt implica'i n eneza 6/M secundar$ ce
apare ncepnd cu al doilea contact cu antienul declanator
/M au o durat de via' mai scurt$ de 8H zile. <le snt alctuite din cinci
structuri asemantoare celei apar'innd moleculei de /Y$ +ind$ deci$ un
pentamer. <le snt implicate n 6/M primar$ declanat la primul contact cu
3G
antienul i snt cele mai active imunolo0uline n activarea complementului pe
calea clasic. Au ac'iune alutinant asupra 0acteriilor i virusurilor.
/<:
prezent n ser n cantit'i (oarte mici.
implicat n declanarea reac'iilor de ,ipersensi0ilitate imediat
/A se sete n ser su0 (orm de monomeri$ sau este secretat la nivelul
mucoaselor sau n lapte su0 (orm de dimeri. Au (ost identi+cate dou su0tipuri
de /A$ ce di(er prin structura lan'urilor Q (Q8 i Q2).
/D este prezent n ser cantit in cantit'i (oarte mici. <a apare (recvent asociat
cu /M$ ndeplinind (unc'ia de receptor pentru antien
%$$.sistemul complement
Sistemul complement: este un comple% de proteine serice cu activitate
enzimatic$ citolitic i relatoare care prin activare produc c,emota%ie$
opsonizare i citoliz a aen'ilor patoeni. 5omplementul este alctuit din 2?-AH
de enzime ce snt activate n cascad$ rolurile Bucate de acest sistem n cadrul
6/M +ind cel de a +naliza ac'iunea anticorpilor asupra antienelor i cel de a
realiza recunoaterea nespeci+c a structurilor nonsel($ aceste roluri putnd +
ndeplinite +e separat$ +e complementar.
n mediile lic,idiene$ +ecare (actor al complementului se sete su0 (orm
inactiv
Mn (actor al complementului odat activat$ va avea drept su0strat un alt (actor n
(orm inactiv$ pe care l va activa$ 5onversia enzimatic a +ecrui (actor inactiv
n (orma sa activ este un proces de proteoliz limitat$ n urma cruia rezult
dou (ramente: un (rament mic$ notat cu NaO i unul mare$ notat cu N0O$ acesta
din urm putnd (unc'iona n cadrul cascadei de activare a complementului ca
enzim$ sau co(actor (+%ator pe mem0rana celular a unui alt (actor).
#actorii complementului snt nota'i de la 58 la 5G.
Activarea complementului se (ace pe mai multe ci. /ni'ial au (ost descrise calea
clasic i cea altern de activare$ care snt distincte dar au o secven' +nal
comun. <venimentul central al cascadei complementului este constituit de
conversia (actorului inactiv 5A n (orma activ 5A0$ su0 ac'iunea 5A-
convertazelor.
5alea clasic de activare este declanat episodic de prezen'a comple%elor
imune ataate mem0ranelor celulare i reprezint o ac,izi'ie +loenetic de dat
mai recent
A treia cale de activare a complementului (ME9 pat,_aT ! mannose binding
lectin path'a() a (ost recent descoperit. ME9 este o protein plasmatic din
(amilia collectinelor$ ce (unc'ioneaz ca o lectin$ +ind sinura apt s activeze
complementul.
Aen'ii patoeni capa0ili de a se lea de ME9 snt Salmonella$ 1eisseria$ 5andida
al0icans. 6eac'iile complementului se deruleaz numai pe supra(a'a
mem0ranelor 'int i nu pe cele sel($ aceastea +ind proteBate anticomplement
prin trei cateorii de receptori
282.6/5
6spunsul imun celular (6/5) este implicat n trei condi'ii patoloice maBore:
eliminarea celulelor in(ectate cu microoranisme cu ,a0itat sau dezvoltare
intracelular (mico0acterii$ virusuri$ leionella$ 0rucela)$ suprave,erea i
aprarea antitumoral i reBetul re(elor de 'esuturi sau orane. "rin urmare$ 6/5
Boac un rol esen'ial n recunoaterea i ndeprtarea prin distruere a dou
cateorii celulare: celulele proprii sau sel( devenite antienice (prin in(ectare sau
deenerare malin) i celulele strine$ nonsel($ provenite prin transplant de
'esuturi sau orane.
LH
"entru realizarea acestor (unc'ii$ este necesar participarea a trei tipuri de
lim(ocite e(ectoare: lim(ocitele T citoto%ice (9T5)$ celulele natural Siller (1U) i
celulele Siller (U). Aceste tipuri celulare recunosc celulele 'int prin mecanisme
di(erite$ dar le distru prin mecanism asemntor$ de citoto%icitate e%tracelular
9im(ocitele T citoto%ice e%prim pe supra(a'a lor cele trei cateorii de receptori:
cei de recunoatere antienic$ T56$ care sta0ilesc cone%iuni att cu antienul
e%pus$ ct i cu molecula prezentatoare a acestuia$ M=5/ sau M=5//. din aceast
cateorie (ac parte receptorii 5DA i 5DD.
A doua cateorie de receptori e reprezentat de cei cu rol accesor n activarea
9T5$ (5D2?$ 5D2D i 5DC?).
5ea de a treia cateorie e reprezentat de receptorii de adeziune intercelular$
reprezentat de 5D2. n (unc'ie de anumite particularit'i (enotipice i
(unc'ionale$ 9T5 se mpart n dou cateorii: 9T5 5DD4$care reprezint peste GHR
din totalul acestora i au rol distructiv i 9T5 5DC4$ care reprezint su0 8HR din
totalul 9T5 i ndeplinesc rol preponderent imunorelator$ intervenind n matura'ia
lim(ocitelor T=p care devin 9T=8.
9a 0aza derulrii 6/5 st participarea celulelor T cu memorie$ dei nu toate
lim(ocitele ce se resesc n snele peri(eric apar'in acestei cateorii. Ast(el$
e%ist lim(ocite T naive$ inocente$ (r nici un contact anterior cu antienul
pentru care au (ost proramate enetic i lim(ocite T cu memorie$ ce au avut
contacte repetate cu antienul pentru care au (ost proramate. 5ele dou
cateorii de lim(ocite T se di(eren'iaz (enotipic$ parcur ci di(erite i au
distri0u'ii (,omin-uri) di(erite la nivelul oranelor lim(oide secundare
"rima etap a 6/5 este cea de induc'ie$ ini'iat n (ocarul antienic$ dar derulat
mai ales intraanlionar. 9a +nele acestei etape$ are loc apari'ia celulelor T cu
memorie i creterea e%presiei receptorilor de adeziune intercelular la nivelul
celulelor din acest (ocar. Aceast prim etap este ini'iat de ptrunderea
intratisular a antienului$ care este captat imunoloic nespeci+c de 5"A$ n
special de macro(aele rezidente la acest nivel. > parte dintre macro(aele
implicate n acest proces rmn la nivel intratisular$ participnd la declanarea
unui proces in-amator. > alt parte dintre macro(ae mireaz pe calea
lim(aticelor a(erente i transport intraanlionar antienul captat la nivel tisular.
5ea de a doua etap a 6/5 se deruleaz intratisular i implic aderarea
lim(ocitelor T cu memorie la endoteliul vascular$ traversarea prin diapedez a
capilarului i cooperarea la nivelul (ocarului antienic cu macro(aele rmase
intratisular. 6ezultatul +nal al acestei etape este distruerea complet i
ndeprtarea antienului declanator.5elulele implicate n aceat etap snt:
macro(aele$ lim(ocitele T=8$ lim(ocitele T5 5DD4 i lim(ocitele T5 5DC4.
RE=A?
%$&.structura unctionala a neronului
1e(ronul reprezinta unitatea mor(o-(unctionala a rinic,iului. "arenc,imul renal
este compus din numerosi ne(roni$ +ecare din acestia posedand o iriatie
sanuina proprie. Mrina se (ormeaza ca urmare a activitatii acestor ne(roni$ iar
(unctia renala$ in totalitate$ poate + considerata ca o suma a (unctiei e%ercitate
de cele apro%imativ 2 milioane de ne(roni.
1e(ronul este compus din corpusculul renal (lomerulul Malpi,i) ce este implicat
in (ormarea urinei si este situat in zona corticala si dintr-un conduct ! tu0ul
urini(er$ ce intervine in concentrarea +nala a urinei si serveste drept conduct
e%cretor$ +ind situat atat in corticala cat si in medulara. Aceste doua componente
provin la em0rion din primordii separate$ conectandu-se apoi in mod secundar.
L8
Ylomerulul$ la randul sau este alcatuit din capsula Eo_mann (e%tremitatea
s(erica inc,isa a tu0ului) si dintr-un ,em de capilare.
Ylomerulul este su0impartit in A-? lo0uli$ (ormati dintr-un numar de anse capilare
tri0utare aceleiasi arteriole a(erente si separate printr-un tesut intercapilar$
denumit tesut mesanial. Supra(ata totala a endoteliului capilarelor este de
apro%imativ 8$? m2. <ste interesant de su0liniat (aptul ca (oita interna a capsulei
Eo_man urmareste ansele capilare de care adera intim
"ortiunea tu0ulara a ne(ronului incepe la nivelul lomerulului si su(era mai multe
cuduri in aceasta reiune$ merand in eneral spre e%terior$ in corte%. Tu0ul se
indreapta apoi si co0oara in linie dreapta spre su0stanta medulara. "ortiunea
sinuoasa si prima parte a portiunii descendente constituie tu0ul pro%imal$ cu o
lunime de 8C mm (82-8C mm). "eretele saueste (ormat dintr-un epi-teliu
unistrati+cat$ cu celule in (orma de piramida trunc,iata$ avand un nucleu
voluminos asezat in pozitie 0azala. 5aracteristic acestor celule este prezenta
marinii jin periek la nivelul polului luminal
/n partea terminala a portiunii descendente$ tu0ul devine e%trem de su0tire.
Aceasta portiune poarta numele de sementul su0tire al ansei =enle. Dupa o
cudura$ realizand aspectul in jac de paru$ tu0ul urca indarat spre lomerulul sau$
in corticala$ realizand ansa =enle$ cu o lunime de 2H-22 mm. De-a lunul
sementului ascendent tu0ul se inroasa din nou$ continuandu-se cu o portiune
canaliculara care prezinta cateva sinuozitati$ pe o lunime de ? mm$ denumita
tu0 distal$ si care se varsa in sistemul canalelor colectoare (2H mm). Aceste
canale mer in linie dreapta prin su0stanta medulara$ colecteaza continutul
rezultat in urma activitatii mai multor ne(roni si se desc,id apoi la nivelul
papilelor renale. "eretele tu0ului contort distal este (ormat$ de asemenea$ din
epiteliu
monostrati+cat$ ale carui celule cilindrice prezinta miorovili$ (ara marine jin
periek.
%$(.aparatul .u1ta'lomerular anatomie si rol
Alcatuit din celule .u1ta'lomerulare si macula densa. Este situat in zona
+ilului fecarui 'lomerul.
0elulele musculare din tunica medie a arteriolei aerente la contactul cu
macula densa sunt 'lo-uloase, af-rilare si contin 'ranule de renina cu
rol de -aroreceptori ce creste productia de renina cand nu sunt
destinse.
?a contactul portiune 'roasa ansa +enle7tu- contort distal celulele sunt
mai inalte, aparatul 'ol'i spre arteriole si au rol in controlul unctiei
neronului, dau semnale de eed -ac9 pentru arteriolele aerenta si
eerenta.
%$).circulatia renala rol,metode de determinare,valori normale
Reprezinta %)E din de-itul cardiac de repaus;$,%)l san'e3min.
@istri-utie neuniorma 9!E in corticala si $! in medulara A 9Ein
medulara e1terna si $ in medulara internaB.
8lu1ul scazut al medularei interna nu spala activitatea osmotica de la
acest nivel, ci mentine anomalia osmotica.
@e-itul san'uin renal se determina prin metode directe de-imetrie pe
vase sau indirecte principiul 8ic9 se calculeaza cantitatea de
su-stanta trasoare preluata de rinic+i in unitatea de timp si impartind7o
la dierenta arterio venoasa. Se oloseste acid paraamino+ipuric sau
diodrast, dozandu7se concentratia lor plasmatica si urinara. 0oefcientul
acestor su-stante de e1tractie este oarte mare la concentratii san'uine
reduse 7K !,9! pt PAC si !,8) pt diodrast.
L2
8lu1ul plasmatic renal este clearance7ul renal. 0learencul ; concentratia
urinara a su-st : volumul de urina intr un minut 3 concentratia
plasmatica a su-stantei
@e-itul renal ; *&!ml3min
8lu1ul renal plasmatic;,!!ml3min
8lu1ul san'uin renal;$%,&ml3min
Ltilizarea 'azelor radioactive a permis calculul /u1ului san'uin in
corticala, )ml3' de tesut3min in medulara e1terna 7 K,ml'3tes3min si in
med interna su- !,%.
Timpul de trecere al eritrocitului este de %,)s in corticala si %,,, s in
medulara.
%$*.consumul de o1i'en in rinic+i
San'ele venos renal are o cantitate de o1i'en mai mare decat in alte
tesuturi.
@ierenta arterio venoasa; $,(7$,, mlE este constanta in variatii ale
/u1ului san'uin.
l consum mediu de o1i'en; (!! u mol3min3$!!' ; (78E din
consumul total de o1i'en al or'anismului
l la de-iti san'uin renal; *7,ml3min3$!!', consum zilnic $87
%$ml3o%3min pentru cei doi rinic+i
l la nivelul corticalei consumul este mai mare datorita transportului
activ al sodiului.
l consumul rinic+iului de o1i'en este crescut datorita
meta-olismului tisular.
l
%$,.autore'larea circulatiei renale
la presiuni de peruzie de 8!7%!!mmC' constanta a de-itului renal
datorita mecanismelor mio'ene si maculei densa.
Se o-serva in corticala autore'larea circulatiei renale. In medulara
variaza cu presiunea arteriale. Se pertur-a in eort fzic prin cresterea
sodiului, se produce vasoconstrictie pe arteriola aerenta, scade de-itul
san'uin, iar su-stantele pireto'ene cresc.
%$8.structura mem-ranei fltrante 'lomerulare si celulele mezan'iale
7 celule endoteliale, epit capsulei 6oMmann
7celulele endoteliale au mii de pori mici; enestre
7pe supraata celulelor se a/a un strat de 'licoproteine polianionice ce
ormeaza 'licocali1ul.
#em-rana -azala are & straturi2
7 central, dens; lamina densa, este mar'init de zonele su-endoteliale si
su-epiteliale lamina rara interna si e1terna. Alcatuit din f-re de
cola'en si proteine plasmatice incarcate ne'ativ ce sunt respinse si
ormeaza spatii lar'i prin care lic+idul fltreaza mai usor.
7epiteliul capsulei 6oMmann la supraata 'lomerulara are prelun'iri
care delimiteaza ante pori la stratul e1tern al mem-ranei -azale
7 intre pediculi pe supraata mem-ranei -azale se 'aseste diara'ma de
anta care are rol de atasare si mentinere a pediculilor. Se 'asesc de
asemenea retele de canale intercelulare prin care fltratul a.un'e in
spatiul 6oMmann. ?a acest nivel e1ista 'licoproteine cu aci sialic.
0elulele mezen'iale ; intre capatul a1ial, sunt incon.urate de su-st
dund si mem-rana a1iala. Retea e1tinsa de microflamente alcatuita din
ala actinina si miozina; mezan'iu. #icroflamentele au rol de a in+i-a
distensia peretelui cap secundara si de a creste presiunea intracapilara
la )!mmC'.
LA
0elulele mezen'iale a'ociteaza comple1e imune si au microflamente
contractile ce produc cito9ine.
%$9.prezentati actorii de care depinde fltrarea 'lomerulara
Mem0rana +ltranta lomerulara. 5aracteristicile anatomice si circulatia
lomerulara lasa sa se intrevada rolul important al (actorilor ,emodinamici in
realizarea ultra+ltrarii unei mari cantitati de plasma prin peretele capilarului
lomerular. Avand proprietati asemanatoare mem0ranelor poroase arti+ciale
(colodiu$ celo(an$ portelan etc.)$ peretele capilarului lomerular se comporta ca o
mem0rana traversata de nenumarati pori
- Permea-ilitatea mem-ranei fltrante
- Supra(ata de +ltrare
- Yradientul de presiune ,idrostatica si coloid osmotica de o parte si de alta
a mem0ranei lomerulare
5ap lom ! permea0ilitate ( mare si selectivitate mare in privinta marimii
moleculei.
/nsulina are aceeasi permea0ilitate ca ,2H.
"roteine cu u KAH HHH dal ! au permeailitate de H$?. 5ele cu u mai mare de
AH.HHH au permea0ilitate H$H?.
#oarte permea0i pt su0st dizolvate in plasma si impermea0ile pt proteine
plasmatice ! al0umina trece in catitati reduse datorita incarcarii neative si
respinerii electrostatice de catre proteinele plasmatice.
=0 este mai putin incarcata neativ si trans(erata in urina primara in
proportie de ?R din concentratia plasmatica.
5ompozitia +ltratului lomerular este ac cu a lic,idului ce +ltreaza la
capatul arterial al capilarului (plasma ce nu are proteine in cantitati
crescute).
5and se pierd sialoproteinele apare al0uminurie (ara modi+carea
dimensiunii porilor ! lomerulo ne(rita si ne(roza lipoidica.
Supraata de fltrare depinde de numarul ne(ronilor 8$2-8$?m2 pentru
cei doi rinic,i.
- Variatii prin contractia celulelor mezaniale si alterarea podocitelor care
pot acoperi portiuni din mem0rana 0azala$ ce determina scaderea
supra(etei de +ltrare
- Aniotensina //$AD= determina contractia celulelor mezaniale si scad
supra(ata +ltranta$ dar pot + rela%ate de "A1
- /n scleroza renala$ ne(rectomie partiala ! supra(ata se reduce datorita
distruerii unor ne(roni
Presiunea eectiva de fltare presiunea ,idrostatica
intralomerularaK3H-LHR din presiunea arteriala sistemicaK3Hmm=
7presiunea din capsula Eo_mannK8Dmm=
7presiunea 5oK2Dmm= la intrarea in capilar si creste la A3 mm= la
iesirea din capilar ! pe masura ce ,2o intravascular ultra+ltreaza$
presiunea 5o medieKA2 mm=.
7presiunea co in capsula Eo_mann este H.
Ionii nu sunt in concentratii identice, urina primara are cu )E mai
multi anioni si cu )E mai putini cationi ec,ili0rul Donnan.
%%!.presiunea de fltrare 'lomerulara
#iltrarea lomerular este rezultatul Bocului dintre (or'ele (avora0ile +ltrrii i
ale celor
opozante. Sinura (or' (avora0il +ltrrii este presiunea hidrostatic a
s&nelui din capilarele
lomerurale (LH mm=)$ +ind cea mai mare presiune ,idrostatic din ntre
teritoriul capilar al
LC
oranismului$ datorit emeren'ei aproape perpendiculare$ din aort$ arterele
renale$ c&t i
rami+crii dup un traiect scurt a acestora la care se adau cali0rul mai
redus al arteriolei e(erente dec&t al arteriolei a(erente. #or'ele opozante
+ltrrii sunt presiunea coloid)osmotic a proteinelor
plasmei (2? mm=) i presiunea capsular (8? mm=). *resiunea efectiv de
ltrare va + tocmai
di(eren'a dintre presiunea ,idrostatic i suma presiunilor opozante.presiunea
e(ectiva de +ltrareKAHmm=
%%$.prezentati rezultatul ultrafltrarii
Proteinele retinute; polianioni. Anionii sunt respinsi si se retin cationii
; ec+ili-rul @onnan
%%%.de-itul fltrarii 'lomerulare
$%)ml3min la -ar-ati si $$!ml3min la emei.
Este cantitatea de fltrat care se ormeaza in fecare minut in toti
neronii.
De0itul urinii primare depinde direct de mrimea presiunii e(ective de +ltrare$
iar aceasta
depinde de mrimea presiunii s&nelui la nivelul capilarelor lomerurale.
5&nd presiunea
,idrostatic din capilare scade su0 CH mm=$ (ormarea urinii nceteaz i se
instaleaz anuria.
5&nd presiunea ,idrostatic din capilare crete$ are loc o cretere al de0itului
urinii primare. Dar$
+ecare ne(ron posed mecanisme de autoreglare a presiunii ,idrostatice
capilare$ men'in&nd-o$
constant n Bur de LH mm=$ c,iar dac tensiunea n arterele renale variaz
de la DH la 2HH mm=.
Aceast relativ autonomie a presiunii de +ltrare se realizeaz prin constric'ia
sau dilata'ia
arteriolelor a(erente i e(erente. ast(el$ constric'ia arteriolelor a(erente i
dilatatrea celor e(erente
scade presiunea e(ectiv de +ltrare$ iar constric'ia arteriolelor e(erente i
dilata'ia celor a(erente
crete presiunea e(ectiv de +ltrare.
Se masoara indirect prin clearance ul la inulina.
0oefcientul de fltrare; cati ml de ultrafltrat se produc3min la
presiunea eectiva de fltrare $mmC';$%,)ml3min
0aract permea-ilitatii fltrului renal depinde de supraata de fltrare si
este in/uentat de celulele mezan'iale.
Rata fltratului 'lomerular este $%)ml3min
8ractia de fltrare este procentul din /u1ul plasmatic renal ce devine
fltrat 'lomerular.
8actorii care in/uenteaza fltratul 'lomerular sunt reprezentati de 2
7de-itul san'uin renal creste /u1ul renal, creste de-itul fltrarii
'lomerulare si invers.
7variatiile de cali-ru a celor % arteriole aerenta si eerentaN
7arteriola aerenta vasoconstricte , scade de-itul rnal si fltratul
'lomerular se produce anurie, vasodilatatie
7arteriola eerenta vosoconstrcitia determina cresterea presiunii
'lomerulara ceea ce determina cresterea actorului 'lomerular ce are
aect -iazic si tranzitor.
Tensiunea arteriala sistemica 8!7%!!mm C', de-itul variaza putin.
L?
%%&.autore'larea fltrarii 'lomerulare
7 mecanism mio'en2 cresterea -rusca a presiunii arteriale determina
cresterea /u1ului renal,apoi apare contractia musculaturii netede din
arteriola aerenta doatorita intinderii si determina scaderea de-itului
arteriolei eerente.
7eed -ac9 tu-ulo 'lomerular Arol al lic+idului T@B2 mec de eed -ac9
vasodilatator al art aerente si determina un de-it scazut al /u1ului
tu-ular datorita rea-sor-tiei crescuta de sodiu si clor. ?a nivelul ansei
+enle ascendente concentratia scazuta a acestora determina la nivelul
maculei densa un semnal dilatator al arteriolei invecinate si creste
de-itul fltrarii 'lomerulare.
#ec de eed -ac9 vasocons al art eerente duce la concentratii scazute
de sodiu si clor din macula si determina eli-erarea de renina.
An'iotensina % este un vasoconstrictor art eerente si determina
cresterea presiunii intra'lomerulare cu sporirea fltratului.
Lrina poate trece de la nivelul capsulei 6oMmann in sistemul tu-ular
T0P2 urina primara;izotona cu plasma $8!l3%(+
A+ descendenta2 la capat urina +ipertona
A+ 'roasa si T0@2 urina +ipotona
T02 urina +ipotona3+ipertona inunctie de starea de +idratare.
0antitatea de fltrat 'lomerular se diminueaza mult2 $%!!7$)!!ml3%(+.
Tu-ul renal are rol de rea-sor-tieAeconomisire a su-t care au ost
antrenate in fltratul 'lomerularB si de secretie; 'rup de su-st
endo'ene care tre-uie eliminate mai rapid decat poate ace fltratul
'lomerular.
%%(.enomenul diurezei presionale si eectul stimularii simpatice asupra
/u1ului san'uin renal
Eectele stimularii simpatice asupra /u1ului san'uin renal enomen
de scapare. #ecanismele intrinseci de autore'lare sunt mult mai
efciente. @upa o stimulare simpatica -rusca, puternica apare anuria.
@aca stimularea persista cateva ore, presiunea creste sufcient pentru a
contracara eectele vasoconstrictoare simpatice.
@iureaza presionala2 din studiul mecanism de rea-sor-tie tu-ulara
rea-sor-ia nu creste cand creste presiunea, de aceea oricare ar f
cresterea fltratului 'lomerular determina automat si cresterea de-itului
urinar. Acest eect pronuntat al presiunii art asupra de-itului urinar
determina diureza presionala.
%%).rea-sor-tia lic+idului in capilarele peritu-ulare si modifcarile
osmoralitatii urinii de a lun'ul tu-ului urinier
"e+icularea urinii depinde de 'radientul de presiune +idrostatica dintre
capsula 6oMmann A$!mmC'B si -azine A!B. ?a nivelul T0@;,mmC'
Epiteliul tu-ular rea-soar-e 99E din C%O din fltrat di cantitatea
crescuta de electroliti.
Presiunea capilarelor peritu-ulare este de $&mmC', presiunea interst
renala;*mmC' ce duce la un 'radient de presiune pozitiv care se opune
rea-sor-tiei
Acest 'radient este contracarat de presiunea co a plasmei;&%mmC' si
Pco din interstitiu;$)mmC'. Presiunea de rea-sor-tie va f $,mmC'.
Presiunea neta de rea-sor-tie este $!mmC' ce permit rea-sor-tia unor
cantitati crescute de lic+id din tu- in interstitiu.
%%*.rea-sor-tia renala a sodiului
*!!' se fltreaza in %( de ore. Se rea-soar-e activ pe toata lun'imea
neronului cu e1ceptia ram descendente ansa +enle.
L3
In tu-ul contort pro1imal2 transportul activ primar in e1tremitatea
-azala, prin pompa %=a3%O7 'radient puternic ne'ativ; 7,!m"
0ei doi actori determina diuziunea =a din lic+idul tu-ular in celula2
7<radientul de concentratie lic+id tu-ular;$(!mEP3l intracelular si
atractia ionilo de catre 'radient electric de 7,!m".
Transportul activ secundar7co transport2 nu oloseste ener'ie urnizata
direct din ATP. #isc =a activ transport secundar.
7simport =a3'lucoza, =a3aminoacizi, =a3O, 7 'lucoza, aminoacizii stra-at
mem-rana -azolaterala prin diuzie acilitata
7antiport =a3C o parte a =a trece in san'e la sc+m- cu C si O
In tu-ul contort pro1imal se rea-oar-e apro1imativ *)E din =a,
rea-sor-tie o-li'atoei, +ormonoindependenta.
Ansa Cenle7portiunea 'roasa se rea-soar-e pasiv %)E, urmand
a-sor-tia activa de cl, e1ista o pompa activa de clor.
/n T5D si T5 se rea0sor0e D-8HR din 1a +ltrat prin mecanism activ primar -ZAT"-
aza 1a@S$ 1a@= controlat de aldosteron.
/n alcaloza 1a@U
/n acidoza 1a@=
/n alcaloze asociate cu ,ipopotasemie se elimina = -Zacidurie parado%ala !
retinerea U este mai pretioasa decat retinerea =.
%%,.re'larea rea-sor-tiei de natriu
8actori de care depinde2
78iltratul 'lomerular2 vasoconstrictia art renala determina scaderea
disproportionala a natriurezei ata de scaderea fltratului 'lomerular
7'radul de e1pansiune a volumului ?E0 e1ista un ec+ili-ru
tu-ulo'lomerular la nivelul ratei fltratului 'lomerular, dar si un
mecanism ec+ili-rul 'lomerular7 tu-ular ; capacitatea rinic+iului de a
creste rata rea-sor-tiei datorita cresterii incarcarii tu-ulare 7. Previne
supraincarcarea T0@ cand creste rata fltratului 'lomerular.
7cand fltratul 'lomerular este direct proportional cu solicitarea tu-ulara
-alanta 'lomerulara tu-ulara este controlata de Pco si +idrostatismul
din capilarul peritu-ular. 0reste presiunea +idrostatica, ceea ce duce la
scaderea rea-sor-tiei de na. 0and scade presiunea +idrostatica, creste
rea-sor-tia de na.
Aldosteronul creste rea-sor-tia de na in tu-ul contort distal.
In +iperaldosteronism cronic se instaleaza retentia sodata si
e1pansiunea volemica si apoi se revine la nivelul initial al e1cretiei de
=a; enomen de scapare datorita scaderii rea-sor-tiei de =a in T0@.
Rea-sor-tia activa de =a are loc in toate se'mentele neronului distal.
Prosta'landinele determina natriureza> modifcarea rea-sor-tiei distala
Cormonii tiroidieni stimuleaza rea-sor-tia de na si +%! din tcp prin
cresterea permea-ilitatii pt 9.
A@C in e1ces creste rea-sor-tia de +%! in T0@ si T0 si duce la scaderea
=a e1tracelular.
%%8.rea-sor-tia aminoacizilor si proteinelor
Aminoacizii sunt rea-sor-iti activ in T0P. =a este necesar. Reapsor-tia
este aproape completa, urina are $7%E aminoacizi.
T0P asi'ura pe toata lun'imea rea-sor-tia aminoacizilor.
Pentru aminoacizii neutrii & sisteme de transport2 pentru aminoacizii
neutrii
0istinoza;-oala de meta-olism> deect al rea-sor-tiei aminoacizilor
care se pierd prin urina, se elimina Ar',?is, ornitina7Kcalculi de cistina
care se depun la nivel renal, in cristalin si in+i-a rea-sor-tia 'lucozei si
osatilor.
LL
E1ista o competitie intre aminoacizii esentiale si cei neesentiali.
Rea-sor-tia proteinelor;&! ' proteine trec prin fltrul renal in %( ore.
Rea-sor-tia se ace prin pinocitoza la nivelul T0P2 se ataseaza pe
mem-rana celulara ce se inva'ineaza si ormeaza vezicula in interiorul
careia are loc descompunerea proteinelor in aminoacizi care sunt
a-sor-iti.
Proteinuria fziolo'ica;$)!m'3zi. 0reste in eort si sarcina. (!E sunt
al-umine serice, $)E imuno'lo-uline si (!E proteine renale.
%%9.rea-sor-tia ureei
%)7&!' uree3%(+ care se fltreaza si se rea-sor- varia-il prin tu-ii renali.
7(!E se rea-sor- pasiv prin diuziune in tu-ulcontort pro1imal si apoi in
tu-ul contort a.un'and la concentrarea urinii.
7mem-rana tisulara este oarte permea-ila pentru uree, de aceea pe
masura ce +%! se rea-soar-e in tu-i, concentratia ureei creste,
'radientul de concentratie in tu-ul urinier este un lic+id peritu-ular cu
diuziune a ureei in capilar.
0antitatea de uree rea-sor-ita depinde de diureza.
In antidiureza cand se rea-soar-e 99E din fltratul 'lomerular, *!7,!E
din ureea fltrara retrodiuzeaza in san'e> in diureza intensa cu de-it
urinar de %ml3min retrodiuzeaza (!E
Tu-ul contort ultima parte necesita prezenta A@C, la acest nivel ureea
se concentreaza si trece in interstitiu in vecinatatea ansei +enle.
Ansa +enle 'roasa, tu-ul contort distal si tu-ii contorti corticali sunt
impermea-ili pentru uree.
Su- actiunea A@C, C%O din se'mente incipiente ale tu-ului contort
trece in interstitiu ureea cu concentratie de (,) mosm3l in urina primara
se concentreaza pana la (!!7()!mosm3l
Tu-ii contorti medulari in prezenta A@C sunt permea-ili pentru uree si
se produce diuziunea in interstitiu datorita 'radientului de
concentratie.
Se acumuleaza uree in zona papilara unde se -loc+eaza datorita
mecanismului de contracurent din vasa recta> din interstitiu diuzeaza
prin peretii ansei +enle su-tiri ascendente si a.un'e din nou in urina
unde este recirculata pentru a creste osmolaritatea la acest nivel.
Procesul de diuziune este stimulat de molecule specifce notate LT.
0learence ul ureei este de ,)ml3min la de-it urinar;%ml3min. @aca
de-itul urinar creste, retrodiuzia este limitata pana la ,)E din rata
fltrarii 'lomerulare.
%&!.rea-sor-tia 'lucozei
Sistemul de transport activ cu capacitatea limitata. =u se elimina
prinurina, intrea-a cantitate este rea-sor-ita in T0P. ?a polul apical
'lucoza intra in celula prin simport cu =a. ?apolul -azal trece in mediul
intern prin diuziunea acilitata. Procesul este limitat de capacitatea de
rea-sor-tie a tu-ilor e1primata prin transport ma1im de 'lucoza ce
depinde de concentratia plasmatica, de-itul fltrarii 'lomerulare,
capacitatea de rea-sor-tie tu-ulara. 0onc concentratia 'lucozei creste
peste un nivel critic; pra' renal determinat de 'radul de saturare al
carausului 'lucoza apare in urina. 0and 'licemia; 8!7$!!m'E, si
fltrarea 'lomerulara;$%)ml3min in fltrare trec $!!7$%) m'
'lucoza3minut.
Transportul ma1im transtu-ular pentru 'locoza este la
-ar-ati&,)m'3min si la emei &!!.
Pra'ul renal;concentratia plasmatica la care 'lucoza incepe sa se
elimine renal. 0oncentratia pra' 'lucoza;%(!7K&!!m'E. Pra'ul
LD
renal;%!!m'E. Pra'ul renal variaza invers proportional cu de-itul
fltrarii 'lomerulare.
In dia-et za+arat la varstnici cu 'licemie crescuta pra'ul renal creste,
'licozuria nu apare pentru ca fltratul 'lomerular este scazut datorita
an'iosclerozei A'lucoza se rea-soar-e totalB.
Rea-sor-tia; izoosmitica si e urmata de controlul volemic al ultra
fltratului.
%&$.rea-sor-tia clorului
Anion principal care acompaniaza =a.
In T0P rea-sor-tia activa secundara simport =a3O3%0l si rea-sor-tie
pasiva urmeaza =a Adatorita incarcarii sale electriceB
Ansa Cenle se'mentul su-tire diuziune pasiva a 0l spre interstitiu
pentru ec+ili-rarea osmotica cu interstititiu +iperton
7 portiunea 'roasa transport activ secundar> la nivelul mem-ranelor
apicale se a/a un caraus care transporta din lume in celula epiteliala
%0l3=a3O
=a este transportat activ in interstitiu, O diuzeaza pasiv in lumen
mentine diuziunea de potential potential care stimuleaza cresterea
cationilor
0l diuzeaza pasiv in intserstitiu .
#ecanismul co7transport al clorului impreuna cu =a la nivelul
mem-ranelor luminare
%&%.rea-sor-tia apei
Procesul in care su-stantele trec prin mem-rana luminala a celulei
tu-ulare. 0ea mai mare parte are loc la nivelul tu-ului pro1imal;8!E
Ao-li'atorieB, +ormonoindependenta. $9E in tu-ul contort distal si este
acultativa.
Se produce prin mecanisme active ce neceista ATP> cotra 'radientului
de concentratie prin mem-rana -azala, dar si mecanisme pasive2 in
aceasi directia cu miscarea apei solvent dra' > se ace in sensul
'radientului electroc+imic.
In tu-ul pro1imal se produce rea-sor-tia de na, cl, +co&, 9,
+%!,'lucoza, aminoacizi si se secreta +Q, acizi or'anici si 'aze.
@in $8!l urina primara se elimina $7$,)l
C%O se rea-soar-e pe toata lun'imea tu-ului cu e1ceptia se'mentului
de dilutie.
Tu-ul contort pro1imal pasiv prin osmoza urmand =a,0l;*)E 7
rea-sor-tie o-li'atorie
In ansa +enle descdendenta;$)E
In T0@ si T0;$9E7 acultativ su- in/uenta A@C
Rinic+iul poate concentra urina pana la $(!!mosm3l in %(+ se e1creta
apro1imativ *!!mosm de micromolecule2uree,acid uric,osati.
0antitatea minima de apa pt e1cretia lor in concentratie ma1ima este
)!! ml urina3zi;urina o-li'atorie, c+iar si in a-senta aportului +idric.
T0P;urina izotona
In ansa +enle ascendenta mecanismul de concetrare a urinei
impermea-ila pt apa
In ansa +enle se'ment 'ros adaptat pentru transport activ de na si cl
din lumen tu- in interstitiu. Impermea-ilitate pt +%!, uree care raman in
tu-.
T0@ e1tensie a se'm 'ros al ansei +enle. Imperma-ilitate pt +%!>
dilutia lic+idului tu-ular.
A@<7 +%o din T0 trece in interstitiu2 urina se concentreaza, presiunea
osmolarica se e'alizeaza cu cea a lic+idului interstitial.
LG
In antidiureza ma1ima se rea-s 99,,E din +%! fltrara, iar concentratia
urinii fnale este intre $%!!7$(!!mosm3l.
0and A@C lipseste, cel T0 ; impermea-ilitate pt +%o.
?ic+idul +ipoton din se' 'ros ansa +enle din care celulele T0@ si T0
continua sa sustra'a =a, a.un'e la concentratia fnala de &!7(!mosm3l>
rea-sor-tia intensa a solvitilor in se'm distale ale neronului cu
pastrarea +%o in tu-i reprezinta mecanismul renal de e1cretie a urinii
diluate.
%&&.mecanismul de dilutie al urinii
%&(.mecanismul de concentrare al urinii
%&).diureza apoasa si osmotica
@iureza apoasa in'erarea in scurt timp a unor cantitati crescute de
lic+ide +ipotone A$7%lB determina o scadere a rea-sor-tiei +%o dupa $)
minute> eect ma1im;()min cand /u1ul urinar;$%7$)ml3min cu
presiunea osmotica scazuta ce determina diureza apoasa.
?ic+idele a-sor-ite determina scaderea presiunii osmotice cu $!mosm3l
in plasma ce determina in+i-area secretiei de A@C. 0antitatea crescuta
de lic+id +ipoton determina tumeferea si aparitia simptomelor de
into1icatie cu +%o2convulsii,coma.
@iureza osmotica su-stantele micromoleculare care nu sunt
rea-sor-ite in T0P pe masura ce volumul urinii primare nu se
micsoreaza, se concentreaza si prin presiunea osmotica pe care o
e1ercita retin +%o in tu-.
Retentie +%o in T0P scade 'radientul de concentratie al =a din lumenul
tu-ular si celula tu-ular, se in+i-a rea-sor-tia.
Ansa +enle volum crescut de lic+id izotonic
T07 prezenta unei cantitati crescute de su-stante ce nu au ost
rea-sor-ite determina scaderea rea-sor-tiei de apa si duce la
eliminarea unei cantitati crescute de urina;diureza
Rea-sor-tia in T0P este scazuta ata de diureza apoasa cand
rea-sor-tia este normala in T0P.
%&*.e1plorarea rinic+iului ct de concentrare
Pentru a cuantifca pierderea3casti'ul de apa prin e1cretia urinii
concentratiei3diluate7 clearance7ul apei li-ere reprezinta dierenta
dintre volumul urinar si clearanceul osmolar.
0learance7ul osmolar;)ml3min
0and urina ;izoosm cu plasma cl7ul osmotic este e'al cu volumul urinar
si rinic+iul nu concentreaza si nu dilueaza.
0and cl7ul apei este pozitiv, urina este +ipotona. 0and e ne'ativ, urina e
+ipertona. 0and e !, urina izotona cu plasma.
%&,.rolul rinic+iului in controlul osmolaritatii
9!E din osmolaritatea ?E0 se datoreaza =a, iar 'lucoza si ureea
reprezinta &E.
aBsistemul osmoreceptor7+ormon antidiuretic mecanism de eed-ac9.
0onserva +%o si eimina =a si alte su-stante osmotic active, corecteaza
osmolaritatea ?E0
7se determina eli-erarea de A@C
7creste osmolaritatea cu $E.
-Bmecanismul setei ; senzatia de sete apare la cresteri ale presiunii
osmotice peste %8!mosm3l
controlul setei aria preoptica, sunt stimulate de orice actori ce
determina des+idratarea intracelulara2 cresterea =a in ?E0, pierderi de
O.
DH
#ecanismul setei este stimulat de an'iotensina ii, des+idratarea
mucoasei oroarin'iene.
?a cresterea concentratiei =a cu %meP3l este amorsat mecanismul de
in'estie a apei2 acea persoana a atins un nivel de sete pentru a activa
eortul motor necesar pentru a -ea;pra'ul setei.
0onsumul de lic+ide pana la starea de satietate, corespunzator
normalitatii osmolaritatii ?E0 necesita$)7() min pt ec+ili-rul osmotic.
cB apetitul pentru sare mentine concentratia e1tracelulara sodata
normal si volumul ?E0. #ecanismul este asemanator cel al setei2 setea
apare imediat, iar apetitul pentru sare dupa cateva ore.
8actorii care dau acest mecanism sunt concentratia scazuta de na in ?E0
si insufcienta circulatorie2 +ipotensiune si +ipovolemia.
Se maniesta in -oala Adisson.
0entrii implicati2 re'iunea Ava"& din creier2re'iunea antero ventrala a
ventriculului III, centrul setei Aneuroni de la acest nivel sunt
osmoreceptori ce trimit impulsuri nucleilor supraoptici pentru a controla
A@CB.
%&8.rolul rinic+iului in controlul izovolemiei
"olemia controleaza tensiunea arteriala care actioneaza asupra
rinic+ilor. 0resterea volemiei determina unde-it cardiac crescut si
tensiune arteriala crescuta, dar si cresterea diurezei.
Scaderea volemiei determina scaderea de-itului cardiac si a tensiunii
arteriale ceea ce duce la retinerea de lic+ide la nivel renal, ce readuc la
normal volemia.
Re/e1ul de volum2 cresterea tensiunii arteriale determina tensionarea
-aroreceptorilor arteriali si a alotor receptori de intindere din zonele de
.oasa presiune, determina in+i-are re/e1a a sistemului nervos,
vasodilatatie.
%&9.rolul rinic+iului in re'larea tensiunii arteriale
re'larea tens art rapide2 prin -aroreceptori,mec isc+emic S=0,
c+emoreceptori
pe termen mediu minute ore2 mec vasocons al sist RAA. Stress7
rela1area vaselor transerul -idirectional de lic+id prin peretele capilar
in si dinspre ar-orele circ pt rea.ustarea volumului san'uin.
7re'larea pe termen lun'2 7 mecanismul R7lic+ide e1tracel cand
volumul ?E0 creste, presiunea art creste si determina diureza si
natriureza presionala. 0and PA;)!mm+' diureza e !. 0and e $!!,
diureza e normala, cand e %!! creste de *78 ori ata de normal.
@aca presiunea arteriala creste la $)! de-itul renal de apa si sare
creste ata de aport astel incat presiunea arteriale va scadea pana
a.un'e la punctul de ec+ili-ru. @aca presiunea arteriala scade mult su-
punctul de ec+ili-ru, aportul de apa si sare creste eliminarile. Aceasta
revenire a presinii arteriale intotdeauna la punctul de ec+ili-ru;
principiul efcientei nelimitate in controlul presiunii arteriale de catre
mec R7lic+id e1tracelular.
Este imposi-il de modifcat pe termen lun' nivelul presiunii medii catre
o noua valoare ara a modifca unul3am-ii actori ai presiunii arteriale2
fe aport +idrosalin, fe 'radul translatiei eliminarii renale de7a lun'ul
a1ei de presiune ce determina o noua valoare unde cele % drepte s7au
intersectat.
%(!.secretia 9
OQ fltrati in urina primara sunt rea-sor-iti activ in T0P, rea-sor-tia
continua si in ansa +enle.
D8
In tcd si in tc secretia OQ> cantitatea secretata e'aleaza cantitatea
in'erata;()7$!!mEP3zi
Aportul alimentar;)!7$!! mEP3l se ace in % etape2
$.captarea OQ din lic+idul peritu-ular dat =a3O ATP7azei din mem-rana
-azolaterala.
%.diuziunea 9 idn celula in lic+idul tu-ular2 cu cat =a creste in T0@ cu
atat creste secretia de O AOaliureza dupa administrarea de diureticeB.
Potentialul in lumenul tu-ular;7)!mv se datoreaza mi'rarii de =a
intracelular cu modifcarea potentialului de actiune.
0ontrolul secretiei de OQ
7 aldosteron
7aport +idric
7scaderea e1cretiei de O
A@C dependent induse de modifcarile osmolaritatii mediului intern
determina tul-urari ale ec+ili-rului acido -azic, acodoza meta-olica sau
resp acuta determina scaderea -rusca a secretiei de O. Prin trecerea din
cel tu- in ?ec scade 9aliureza. Alcaloza meta-olica sau resp acuta
determina creterea -rusca a 9aliurezei.
%($.mecanismul umoral de control al activitatii renale
%(%.mecanismul intrinsec de autore'lare in mentinerea constanta a
de-itului san'uin renal
%(&.re/e1ul intrinsec de autore'lare in mentinerea constanta a
de-itului san'uin renal
E0CI?I6RL? A0I@O 6A5I0
%((.sisteme tampon e1tracelulare
Cco& ormat din C%0o& si =a%0o&, sistemul osatilor osat
monosodic3osat disodic si sistemul proteinatilo2 proteina acida si
proteiat de natriu, sistemul +emo'lo-itilor;8!E din sist tampon
e1traccelular non-icar-onati. E1ista % sisteme tampn C-2 C- redusa
Apunti saline ormate din CB3+emo'lo-inat de O si o1i C-3O1i C- de O. O
persoana cu anemie severa are capacitatea R de aparare antiacida.
%().sisteme tampon intracelulare
0ea 7 contin ca -aza con.u'ata o sare de O. Sistemul proteinatilor si
sistemul osatilor.
Sist tampon intervin instantaneu in reactie dar se consuma rapid.
0riterii de efcienta ale sist tampon2 toate sist tampon se consuma
proportional in reactie; principiul izo+idriei.
$.cantitatea sist osatilor7 %mEP3l
7 sist proteinatilor %,)7&'3dl
%.valoarea p9 ului2 cu cat 9 este mai apropiat de p+ mediu intern
sistemul tampon e mai efcient. 8osatii au p9 *,8, proteinatii au p9
varia-il *,)7$!,$%.
&.capacitatea de eliminare3to1icitatea prod rezultat in urma tamponarii.
Sistemul -icar-onatilor
0oefcientul de solu-ilitate este $,% mEP3l in san'e arterial. P+ ul are
valori normale ,,&,. 0o% componenta respiratorie pt ca valoarea lui
depinde de 'radul de ventilatie.
%(*.rolul tesuturilor in controlul EA6
#i.loace-iolo'ice de control2 piele,os,musc+i,tract 'astro intestinal cu
'l ane1e2 fcat,pancreas e1ocrin,eritrocit,plamani sist respirator, rinic+i
e1cretie de C, aciditate titra-ila, sintetizare si rea-sor-tie de C0O&
sau secretie de +co& in alcaloza, amonio'eneza.
D2
Pielea secretie sudorala elimina C
#usc+ii controleaza ec+ili-rul acido -azic prin transmielinizare si prin
le'area de proteine musculare, dar activitatea musculara scade in
a'resivitate acida violenta.
Osul7 matrice protectoare saruri si +idro1iapatita2 =a% co&, ca co& si
9%co&> capacitate crescuta de a prelua C> daca a'resiunea e acida;
cronica7 se produce demielinizare osoasa prin dislocarea ca din matrice.
E1cesul de ca care a.un'e e1tracelular se depune in locuri de e1cretie2
canale, 'l salivare, rinic+i litiaza renala.
Stomac secreta .cl in cel cantit crescuta de +co& care trece in mediul
intern in di'estia 'astrica normala p+ mediu int se alcalinizeaza usor.
In a'resiuni acide creste capacitatea stomacului de a ace secretie
acida. In +ipersecretie 'astrica3pierderi masive de suc 'astric in ulcere
duodenale,varsaturi din stenoza pilorica7 pierderea de acid stimuleaza
secretia celulelor o1intice si duce la alcaloza meta-olica si la scaderea
ractiei de calciu ionizat spasmoflie.
Pancreas e1ocrin secr +idrominearala cu p+ alcalin in cel se
acumuleaza C care trec in mediul int in di'estia intestinala initiala
acidiferea mediului int
7 +ipersecretie si pierdere de lic+id intest prin diaree
8icatul deto1ifca or'anismul7 se prelucreaza ac lactic,metanoletanol
salicilati. In insufcienta +epatica scade capacitatea de aparare anti
acida2 in dia-et scade admisisa de 'lucoza la nivel +epatic , scade
capacitatea de meta-olizare .
Eritrocitul sintetizeaza +co& care a.un'e in plasma. 8i1eaza co%7 comp
acida pe lanturile ala si -eta. Preia C puntile saline astel incat
tamponeaza C
%(,.rolul T<I in controlul EA6
%(8.rolul eritrocitului in controlul EA6
Eritrocitul sintetizeaza +co& care a.un'e in plasma. 8i1eaza co%7 comp
acida pe lanturile ala si -eta. Preia C puntile saline astel incat
tamponeaza C
%(9.rolul plamanilor in controlul ea-
Ciperventilatie scazuta de co% alcaloza dar intervin c+emoreceptorii
centrali scade ventilatia> +iperventilatia apre in2 e1punere la
temperaturi crescute, altitudine mare, eort fzic, into1icatia centrului
resp cu aspirina Ala copiii miciB, ver-alizare multa, crize de isterie, calciu
ionizat mic7 ce duce la risc de spasmoflie si tetanie.
Cipoventilatia se ace in cazul persoanelor cu -oli pulmonare cronice si
tul-urari masive de diuziune2 into1icatie cu -ar-iturice, anestezice> se
poate ace voluntar> se acumuleaza co%; acidoza respiratorie, dar
c+emoreceptorii raspund prompt prin cresterea ventilatiei.
#odifcarea p+ ului datorita modifcarii primare a ventilatiei nu este de
durata. Tul-urarile meta-olice modifca ventilatia prin c+emoreceptori
perierici2 in acidoza meta-olica sunt stimulati ceea ce duce la
+iperventilatie si acidoza dia-etica.
In alcaloza meta-olica> sunt in+i-ati c+emoreceptorii perierici
+ipoventilatia corecteaza p+ ul, dar se acumuleaza co% si
c+emoreceptorii centrali sunt stimulati.
%)!.rolul rinic+iului in controlul ea-
Rinic+iul este principalul care controleaza ea-. Reactia sistem tampon si
respiratorie compensare Aapropierea p+ ului la valori normaleB.
Rinic+iul intervine mai lent, dupa cateva zile, dar unctioneaza pe toata
durata dezec+ili-rului. 0orectia se ace prin p+, care va avea valori
DA
normale. P+ ul urinar variaza (,(78. Secretia de C sint de C0O,
amonio'eneza.
8iltratul 'lomerular contine aceleasi elemente ca plasma2 na, 9 , cl, ca,
+co&, + in aceasi proportie cu plasma urina primara are p+ ,,(.
0antitatea de +co& depinde de r'.
%)$.mecanisme renale de eliminare a protonilor
%)%.secretia si rea-sor-tia renala de C0O&
T0P2 an+idraza car-onica in cel tu-ulara si mem-rana apicala nerocit2
in cel tu-ulara reactie de +idratare a co%. CQ se secreta la acest nivel
secundar activ prin transportorul de na3+ si in masura mai mica prin
pompa de CQ> motorul sc+im-atorului; ATP aza =a3O din polul -azal al
celulei, 'radient de concentratie =a intre celula tu-ulara si lic+id
tu-ular.
C0O& trece din nerocit in mediu intern prin antiport cu cl si simport cu
na. C secretati si C0O fltrat 7 KC%0o& in lic+idul tu-ular. Trece in cel
tu-ulara si reintretine procesul
0o% provine din propriul meta-olism si plasma ceea ce duce la
cooperarea plaman7rinic+i2 acidoza respiratorie. 0o% suplimentar la
rinic+i pt ca sintetizeaza +co&. Se rea-soar-e +co& ormat in celula
tu-ulara renala.
In T0@ si T02 celulele principale secreta C si intercataleAmai rareB
rea-sor- C0O&
Lrina 7K e1cretie prin pompe de C si C3O care introduce O in celula si
scoate C. In lic+id tu-ular reactie sla-a intre C si C0O&.
0elula intercalata sc+im-ator apical C0O&3cl 2 secreta C0O& si
reintroduce cl cand p+ ul creste peste ,,(%.
%)&.aciditatea titra-ila
0antitatea de C le'ati de diverse su-stante A#C(QB care se eli-ereaza
in cazul titrarii> principiul sistemului; sist osatilor.
%)(.amonio'eneza
0ea mai importanta cale de aparare renala.
In T0P2 la nivelul nerocitului se meta-olizeaza 'lutamina. In celula
tu-ulara n+( este secretat in lic+idul tu-ularAneroit pro1imalB unde
ramane su- aceasta orma.
@e7a lun'ul restului neronului tu-i impermea-ili pt n+( astel incat nu
diuzeaza in interstitiu> sin'ura zona permea-ila ; ram 'ros ascendent
ansa +enle An+( trece in medulara,disociaza in n+&QC r de disociere in
ec+ili-ru. =+& diuzeaza in T0, iar CQ sunt secretati cu a.utorul
pompelor in nerocit.
In T0 CQ =C&7K=C(;zona impermea-ila pt +n(. 0u cat se e1tra'e mai
mult n+& in medulara cu atat disociaza n+( mai mult si avorizeaza
eliminarea.
P+;(,)2 rinic+iul e1reta +Q ara acidifere suplimentara7 se pot e1creta
8!!79!!mEP sarcini acide.
%)).anion 'ap
Electoneutralitate2 nr anioni;nr cationi
E1tracelular principal cation; na, m', ca,9
Anioni7 cl principal , +co&, so,po,proteinati.
Anion 'ap ; dierenta intre na si suma cl si +co&.
Anion 'ap ; $! meP3l
"alori normale2 $%;37 ( mEP3l
daca se adau'a +cl anion 'ap nu se modifca.
DC
Acidoza acuta sunt aectate cel tcd si ale tc la acelasi nivel, scade
activitatea atp azei na39. In mem-rana apicala scade permea-ilitatea pt
9. Scade e1cretia de 9 ceea ce duce la +iper9alemie.
%)*.parametri de apreciere ai ea-
p+ mediu int; ,,&)7,,(%
Pco%;(!mmC'
Cco% standard ; cantitatea de +co& plasmatic la temp &, 'rade,
p0o%;(!mmC< si presiune atmoserica ,*!mmC'7 nu tine seama de
variatiile renale ale co% plasmatic. 6icar-onat standard %(7%)meP3l
Rezerva alcalina %( meP3l
6aze totale tampon;)!7));suma tuturor sist tampon
0ompensarea la modifcarea primara a unuia din comp raportului
cealalta se modifca in acelasi sens. "aloarea raportului revine la normal
si compensarea nu este perecta. P+ ul compensat usor dierit de cel
normal. Acidoza meta-olica supraproductie de protoni2 insufcienta
+epatica, renala, inanitie prelun'ita, diaree prelun'ita. Poate f
compensata prin +iperventilatie. Acidoza respiratorie creste co%2
+ipoventilatie o-structie -ronsica, edem, emfzem pulmonar, peruzie
inadecvata plaman.compensarea este renala prin rea-sor-tia de +co&.
Alcaloza meta-olica varsaturi 'astrice in spasm piloric,
+iperaldosteronism. 0ompensarea se ace prin +ipoventilatie.
Alcaloza respiratorie2 scade co% +iperventilatia2 eort,+ipertermie,crize
de isterie, into1icarea centrului respirator cu salicilati. 0ompensarea se
ace renal prin cresterea e1cretiei de +co&.
%),.mecanisme de control in acidoza meta-olica
0auze2 tul-urari 'astro intestinale , into1icatii, meta-olice.
Tipuri2 lactacidemie, cetoacidoza cauza e1o'ena si renala.
0auze2 insufcienta circulatorie2 peruzia or' centrale fcat,rinic+i,
scade o1i'enarea, scade transportul ac lactic de la musc+i la fcat.in soc
cardio'en circulator, insufcienta cardiaca con'estiva, inarct masiv, soc
circulator din septicemie.
7+ipo1emie acuta severa7 anemie, into1icatie cu co
7insufcienta +epatica uneori si in des+idratare cu depletie. Scade
peruzia or'anelor.
0etoacidoza dia-etica in des+idratare2 acumulare de corpi cetonici
sunt meta-olizate TA<, de inanitie cronica, alcoolica inanitieAnu se
+ranesteB si varsaturi, scade vol ?E0;actor de stress si creste secretia
de cortizol si catecolamine ceea ce duce la mo-ilizarea lipidelor din
depozit.
@e cauza e1o'ena 2
aB metanol2 in cursul meta-olizarii
-B salicilati nu au caracter acid, dar in cantitate crescuta determina
doua tipuri de reactii +iperventilatie cu alcaloza respiratorie si
meta-olizare +epaticaAacid o1alicB. 0ele doua duc la dezec+ili-ru
mi1t diver'ent cu p+ nemodifcat.
@e cauza renala2 insufcienta renala acuta3cronica2disunctie
'lomerulara si tu-ulara. Acidoza tu-ulara renala ; incapacitatea cel
tu-ulare de a e1creta protoni.
%)8.mecanisme de control in acidoza respiratorie
Pot f non pulmonare si pulmonare
=on pulmonare2 unctie defcitara a centrului respirator into1icatii cu
anestezice. Alterarea transmisiei nervoase2 miastenia 'ravis,
poliomielita de n renic, -oli musculare2 miopatii distrofce.
D?
Pulmonare2 -oala pulmonara acuta +ipo1emie cu stimularea
c+emoreceptorilor perierici. 6ron+opneumopaia cronica o-structiva
severa. 6oli cronice o-structive astm -ronsic2 o1i'enare insufcienta
co% in e1ces.
%)9.mecanisme de control in alcaloza meta-olica
7 depletie volemica +ipovolemia
7+ipo9alemie primara
7+iperaldosteronism
Cipovolemia activarea sist r a a
7 an'iotensina II si aldosteron activeaza atp aza na39 din mem-rana
latero -azala.
7activarea adrener'ica vasoconstrictoare
Cipo9alemia primara2 creste re-sor-tia de 9 in T0P in mediul intern. Se
rea-soar-e si +co& pt ec+ili-rul electric p+ ul intracelular scade dar
creste rea-sor-tia de +co& din lic+idul tu-ular.
7e1cretie de n+( creste
Situatii particulare2 varsaturi 'astrice, administrare de diuretice de
ansa ce cresc e1cretia de na si +%!, post+ipercapnic, se produce
ventilatie artifciala.
%*!.mecanisme de control in alcaloza respiratorie
0auze pulmonare pneumonie medie em-olie, edem pulmonar
f-roza
0auze e1trapulmonare septicemia +iperventilatie, -oli +epatice
Ameta- pro'esteronuluiB. 0reste concentratia plasmatica de
pro'esteron apare +iperventilatia.
In sarcina de la s primului trimestru prin cresterea pro'esteronului
se instaleaza +iperventilatia.
Cemodializa insufcienta renala initial ac meta-olica compensata
respirator prin +iperventilatie> prin +emodializa se indeparteaza
produsii acizi, dar co% scade pana la acomodare.
7 leziuni cere-rale, +iperventilatie psi+o'ena, altitudine.
%*$.sistemul tampon al +emo'lo-inatilor
Sistemul tampon al ,emolo0inei. =emolo0ina isi poate e%ercita actiunea
tampon in special$ datorita (aptului ca cuprinde ! in partea sa proteica ! multe
resturi de ,istidina. Acesta este un aminoacid cu nucleu imidazolic care$ avand
capacitatea sa accepte si sa cedeze =4 la un atom de azot din ,eterociclul sau$
poate (unctiona ca 0aza sau ca acid.
=emolo0ina intra in constitutia a doua sisteme tampon: (a),emolo0ina acida !
,emolo-0inat de potasiu si (0)o%i,emolo0ina ! o%i,emolo0inat de potasiu.
Atat sistemul care cuprinde ,emolo0ina cat si cel care cuprinde ,emolo0ina
o%idata (o%i,emolo0ina) reprezinta cele mai importante sisteme tampon cu
componente proteice din sane.
Daca mecanismele respiratorii nu pot compensa pertur0arile ec,ili0rului acido-
0azic$ (unctia de e%cretie renala reprezinta urmatorul mecanism de control al p=-
ului mediului intern. 6inic,iul intervine in acest proces atat prin rea0sor0tia si
sinteza 0icar0onatului cat si prin secretia =4$ su0 (orma acizilor nevolatili !
pentru +ecare mol de =4 eliminat prin urina se rea0soar0e sau se sintetizeaza un
mol de =5>A-. #unctia rinic,iului in controlul ec,ili0rului acido-0azic este mai
lenta si depinde in special de valoarea p=-ului sanuin. Deci$ rinic,iul$ in (unctie
de valoarea p=-ului sanuin$ elimina +e e%cesul de acizi$ +e e%cesul de 0aze
SISTE#L? =ER"OS
D3
%*%.componentele unctionale ale sistemului de transport ionic
mem-ranar
%*&.clasifcarea unctionala a canalelor ionice
5analele ionice sunt proteine moleculare celulare care (ormeaza pori de-a lunul
mem0ranei celulare. 5analele prezint 0ariere de permea0ilitate i de
selectivitate$ unele din ele operate c,imic sau electric.Aceste structuri proteice
releaza -u%ul electronic al di(eritilor ioni: sodiu (1a4)$ potasiu (U4)$ calciu (5a4
4) sau clor (5l-)$ prin desc,iderea si inc,iderea porilor. #iecare canal ionic este
selectiv$ permitand de o0icei unui sinur tip de ioni sa-l traverseze. 5analele
ionice sunt prezente in mem0rana tuturor celulelor. <le Boaca un rol central in
+zioloia celululor e%cita0ile cum sunt neuronii.
5analele sunt (acilitatori ai di(uziei$ nein-uentand sensul de trecere a
ionilor$dictat doar de di(erenta de potential electroc,imic al ionului
respectiv(suma a di(erentelor de concentratie si camp electric).Mn canal nu poate
transporta ioni impotriva acestui radient$rol detinut de pompele ionice.
Sunt cunoscute cca ?H de tipuri de canale ionice care eneral se pot clasi+ca in
doua cateorii:canale pasive$caz in care controlul circulatiei ionilor se realizeaza
doar prin dimensiunea porilor in raport cu ale ionilor(e% de dimensiuni
caracteristice pt ionii care circula prin mem0rana 2$?Am*U4si5l- .A$?Am*1a4.
C$?Am*5a 24 ) si canale active$ la care controlul se realizeaza prin modi+cari ale
(ormei si dimensiunii canalelor$ provocate de cele mai multe ori prin procedee
electrice si c,imice( canale controlate in tensiune prin valoarea di(erentei de
potential dintre (etele mem0ranei si canale controlate c,imic prin mediatori
c,imici de tipul acetilcolinei si acidului ama-amino0utiric.)
%*(.canale ionice controlate de volta.
%*).canale ionice controlate de mediator
%**.canale ionice controlate mecanic
%*,.canale ionice ara poarta
%*8.canale ionice de sodiu7distri-utie,rol unctional
5. ionic voltaB-dependent ce permite di(uziunea simpla a sodiului /n celula si$ /n
consecinta$ depolarizarea mem0ranei si declan-sarea potentialului de actiune.
Structural este alcatuit dintr-un sinur lant polipeptidic (2?H-2LH SDa) dispus /n
patru domenii transmem0ranare omoloe. #iecare domeniu este alcatuit din sase
lanturi peptidice transmem0ranare cu structura de tip a-,eli%$ delimitand peretii
canalului. Al patrulea lant a-,eli% a) +ecarui domeniu contine sase reziduuri de
arinina Bucand rolul unui senzor voltaic$ care determina desc,iderea canalului.
Den-sitatea c. de s.$ c,iar /n mem0ranele e%cita0ile$ este de ma%inmum
8HH@nm2$ +ind de circa 8Ho ori mai mica decat cea a canalelor de potasiu. "rintr-
un sinur c$ desc,is tree circa 8HL ioni de sodiu pe secunda. pAntaonistii c. de s.
sunt o serie de anestezice locale (lidocaina)$ kantiaritmice de clasa / A (c,inidina)$
anticonvulsivante (car0amazepin) si diuretice (amilo-rid). V. si canal ionic$ canal
de caldu si canal de potasiu
%*9.canale ionice de calciu
8amilia canalelor de potasiu operate electric este e1trem de diversa. Au
rol in codifcarea intensitatii stimulilor, in recventa si adaptarea
neuronala.
5. ionic care controleaza cea mai mare parte din -u%urile trans-mem0ranare de
calciu$ cu implicatii deose0ite /n declansarea si modularea unei ame lari de
activitati celulare. "roteina c. este (ormata din dnci su0unitati polipeptidice si
licoproteice: al(a$ (8L? SDa)$ al(a$ (8CA SDa)$o0eta (?C SDa)$ ama (?C SDa) si
delta (2L SDa). Su0unitatea al(a$ ar contine c. si situsurile de (os(orilare
modulatoare intracelulare. Se cunosc doua clase mari de c.: receptor-dependente
si voltaB-dependente. S-au de-scris numeroase tipuri si su0tipuri cu caracteristici
si localizari di(erite. Mnul dintre cele mai importante este c. de eli0erare de calciu$
DL
din mem0ranele kreticulului endo-plasmic si kreticu-lului sarcoplasmic (/n celulele
musculare)$ care$ cand este acti-vat$ eli0ereaza cationul /n citosol. Tipurile
principale de c. voltaB-dependente sunt T (T K transient$ rapid)$ 1 (1 K neu-ronal)
si /ndeose0i 9 (9 K lon lastin$ lent) asupra caruia actioneaza pin,i0itorii de
calciu. V. canal ionic$ canal de pota-siu si canal de sodiu
%,!.canale ionice de potasiu
5. ionic care permite iesirea potasiului din celula. Au (ost descrise numeroase
tipuri de c. de p.$ a caror desc,idere depinde pentru unele de voltaB$ iar pentru
altele$ receptor-de-pendente$ de lianzi ca ionii de caldu$ acetilcolina si AT". 5.
dependente de AT" sunt in,i0ate de psul(amide antidia-0etice. pAntaonistii sau
in,i0itorii de c. voltaB-dependente sunt n pantiaritmice de dasa a 888-a si
/ndeose0i de clasele / A si / 5. . Antaonistii de c. receptor-dependente sunt oanti-
isc,emici. pAonistii sau activatorii acestor canale sunt vasodilatatoare (prin
rela%area musc,ilor netezi) si$ in consednta$ utiliza0ili oin tratamentui
p,ipertensiunii arteriale si al pinsu+dentei corona-riene. V. canal ionic$ canal de
caldu$ canal de sodiu.
%,$.pompe de calciu
%,%.potentialul de actiune
POTE=JIA?L? @E A0JIL=E =ELRO=A? APAB
" reprezintG inversarea rapidG Ii complet reversi-ilG a
polaritGFii mem-ranei neuronale, care devine electric
pozitivG la interior Ii electric ne'ativG la e1terior
" azele2
perioada de laten ; !,$ ms
depolarizarea 7 pHnG la Q&! m"
7 in/u1 de =a
Q
S canale rapide volta. depend.
repolarizarea - revenire la valoarea potenFialului de
repaus
7 sistarea in/u1ului ionilor de =a
Q
7 e/u1ul ionilor de O
Q
postpotenFialul pozitiv 7 ATP 7 aza =a
Q
3O
Q
7 resta-ilirea ec+ili-rului ionic
0O=@L0EREA POTE=JIA?L?LI @E A0JIL=E T= 8I6RE?E =ER"OASE
n fbrele nervoase amielinice
depolarizarea unei zone mem-ranare produce
activarea canalelor de =a
Q
adiacente
PA se propa'G 7 secvenFial
7 din aproape Un aproape
7 cu vitezG micG
n fbrele nervoase mielinice
conducerea este saltatorie de la un nod Ranvier la altul
e1plicaFia 7 densitatea oarte mare a canalelor de =a
Q
la nivelul
nodurilor Ranvier
7 teaca de mielinG permea-. V a mem-r. internodale
viteza de conducere depinde de2
'rosimea f-rei nervoase
distanFa dintre nodurile Ranvier
%,&.transmiterea sinaptica c+imica
0?ASI8I0AREA SI=APSE?OR2
c+imice
structura 7 componenta presinapticG
7 anta sinapticG
DD
7 componenta postsinapticG
secvenFa enomenelor transmiterii sinaptice2
invazia -utonului sinaptic de cGtre in/u1ul
nervos
eli-erarea mediatorului Un anta sinapticG
propa'area transsinapticG a in/u1ului
nervos
7 diuziune mediatorului c+imic
7 acFiunea med. c+imic asupra
mem-ranei postsinaptice
7 inactivarea mediatorului c+imic
%,(.transmiterea sinaptica electrica
%,).sinapsa in+i-itorie
Potenialul postsinaptic inhibitor se poate produce prin2
desc+iderea canalelor de 0l
7
ce va induce o ne'ativare
rapidG a interiorului celulei
creIterea conductanFei ionilor de O
Q
spre e1terior, cu
eect +iperpolarizant
activarea unor enzime ce in+i-G uncFii meta-olice
celulare sau cresc numGrul de receptori sinaptici
in+i-itori
%,*.sinapsa e1citatorie
Potenialul postsinaptic excitator se poate realiza prin2
desc+iderea canalelor de =a
Q
ce permit unui numGr
mare de sarcini electrice pozitive sG pGtrundG Un
celulele postsinaptice
reducerea conductanFei canalelor de 0l
7
sau O
Q
sau
am-ele
activarea meta-olismului intern celular va creIte
numGrul receptorilor mem-ranari e1citatori sau va
scGdea numGrul celor in+i-itori
%,,.unitatea motorie
9a om masa muscular este (ormat din cca. 2?H milioane de +0re$ inervate
de cca.
C2H.HHH de motomeuroni. deci un neuron motor din coarnele anterioare ale
mduvei spinrii
controleaz un numr di(erit de +0re musculare. Ast(el unit'ile motorii ale
muc,ilor lo0ilor
oculari$ care e(ectueaz micri +ne (oarte precise pot + (ormate c,iar i
dintr-o sinur +0r
muscular n timp ce la nivelul muc,iului cvadriceps$ care e(ectueaz micri
de (or' i de
amplitudine$ unitatea motorie controleaz sute de +0re musculare. Toate
+0rele musculare inervate de un neuron motor se contract i se rela%eaz n
acelai timp de unde i denumirea de unitate motorie$ care desemneaz acest
ansam0lu.
At&t +0rele musculare c&t i unit'ile motorii rspund leii tot sau nimic.
Muc,iul n
ntreimea sa nu se supune acestei lei$ contrac'ia sa +ind radat prin
recrutarea unui numrmai
DG
mic sau mai mare de unit'i motorii. Aceast intrare n ac'iune a unit'ilor
motorii permite varia'ia
(or'ei dezvoltate de un muc,i a-at n contrac'ie. #or'a se realizeaz +e
variind numrul de unit'i
motorii stimulate la un moment dat (sumaie spaial)$ +e (c&nd s varieze
(recven'a de stimulare a unit'ii motorii respective (sumaia temporal).
Mnit'ile motorii pot + mpr'ite dup criterii meta0olice i (unc'ionale n dou
cateorii:
unele apte s lucreze n condi'ii de aero0ioz i altele mai apte n condi'ii de
anaero0ioz. 9a om
unit'ile motorii apte pentru a lucra n condi'i aero0e con'in bre roii! tonice
(de tip I)$ cu secus
lent. unit'ile motorii adaptate pentru lucrul n condi'ii anaero0e con'in bre
albe! fazice
(de tip IIB$ cu secus rapid.
o unitate motorie este constituit dintr-un neuron i +0rele musculare pe care
acesta le inerveaz$ a-ate n numr invers propor'ional cu radul de precizie al
micrilor e(ectuate de ctre muc,iul respectiv (e.. un neuron vaal inerveaz
p&n la 2HHH de +0re$ un a%on din musculocutanat inerveaz cca. 2HH +0re-la
nivelul 0icepsului$ iar un motoneuron din nervii oculomotori inerveaz 2-C +0re
musculare). Mn muc,i primete inerva'ie motorie din mai multe semente
medulare$ iar un sement medular poate inerva mai mul'i muc,i.
%,8..onctiunea neuromusculara
`onc'iunea neuromuscular (placa motorie) este (ormat din 0utonii terminali
i
rami+ca'iile a%onului motoneuronului pe de o parte i sarcoleme +0rei
musculare$ pe de alt parte.
ntre cele dou componente se a- spa'iul sinaptic de cca. CHH q.
Componenta presinaptic
(0utonul terminal) con'ine vezicule cu acetilcolin$ mediatorul c,imic ce
transmite impulsul nervos motor. Componenta postsinaptic +sarcolema +0rei
musculare) con'ine numeroi receptori speci+ci colinerici de care se +%eaz
acetilcolina$ precum i receptori enzimatici (colinesteraze) ce deradeaz
mediatorul c,imic n vederea unei transmiteri sinaptice normale.
%,9.cuplarea e1citatie7contractie
5uplaBul e%cita'ie-contrac'ie: necesit creterea 5a44 citosolic.
Sursa de 5a44:n vasele mari este intracelular (tu0ii 9).n vasele mici intra- i
e%tracelular (de aceea
0locantele canalelor de 5a44pot reduce in-u%ul de 5a44n aceste +0re$
determin&nd vasodilata'ie).
5uplul e%citatie-contractie-rela%are (<56). Acest proces de cuplare electro-
mecanica de+neste lantul de (enomene care realizeaza leatura (unctionala
dintre sarcolema e%citata prin mesaBul contractil motor si su0stratul contractil
mio+0rilar$ si care consta din: depolarizarea sarcolemei . enerarea si propaarea
unui potential de actiune de-a lunul sarcolemei si al tu0ilor transversi.
eli0erearea calciului din cisternele reticulului sarcoplasmatic (6S) . di(uzarea
sarcoplasmatica a ionilor de calciu si learea lor la receptorii 5-troponinici.
anularea e(ectlui in,i0itor al troponinei / asupra interactiunii acto-miozinice.
interactiunea acto-miozinica cu lisarea +lamentelor su0tiri printre cele roases
scurtarea sarcomerului$ av&nd drept rezultat declansarea$ ntretinerea si
ntreruperea contractiei e%primata prin ciclul jcontractie-rela%areO.
Potentialul de actiune a.uta la desc+iderea canalelor de calciu si vor
determina activarea osorilarii miozinei si initierea contractiei.
GH
#ecanismul contractiei in f-ra musculara neteda necesita calciu
citosolic. Activarea laturilor usoare ale miozinei in prezenta de atp si a
ionilor de ma'neziu. Activarea miozinei va antrena cupla.ul acto7
miozinic cu producerea contractiei musculare
%8!.mecanismul contractiei musculare
5ontractia musculara apare din punct de vedere eneretic cu o succesiune de
reactii producatoare de enerie$ de viteza varia0ila si reversi0ile n timp. Aceasta
succesiune poate + pusa n evidenta urmarind variatiile p=-ului local n timpul
contractiei:
- (aza alcalina$ precontractila$ de semni+catie necunoscuta.
- (aza de acidi+ere$ corespunzatoare scindarii AT".
- (aza de alcalinizare$ corespunzatoare eli0erarii (unctiei 0azice a creatinei prin
scindarea (os(aenului necesar re(acerii AT".
- (aza de acidi+ere$ corespunzatoare cresterii productiei de acid piruvic si acid
lactic$ cu eli0erarea eneriei necesare re(acerii (os(aenului si partial a lucozei.
Din punct de vedere termodinamic$ eneria (urnizata musc,iului tre0uie sa +e
eala cu cea pe care el o eli0ereaza su0 (orma de lucru mecanic$ leaturi (os(at
macroerice si caldura.6andamentul mecanic al musc,iului$ e%primat prin
procentaBul eneriei c,imice a nutrimentelor$ utilizat pentru activitatea
musculara$ aBune p&na la 2H-2?R n conditii optime$ L?-DHR din enerie +ind
eli0erata su0 (orma de caldura.
%8$.relatia lun'ime tensiune a sarcomerului
%8%.meta-olismul muscular
%8&.tipuri de f-re musculare
8i-re rosii de tip $ celule mici cu multe mitocondrii, reticul
sarcoplasmic redus,multe capilare,rata aero-a crescuta, rata 'licolitica
scazuta, rezerve scazute de 'lico'en,ATP7aza miozinica lenta,contractie
lenta.
8i-re o1idative lente 7$3( din dimensiunea f-relor al-e,eect usor spre
moderat .
8i-re al-e de tip % celule oarte mari, putine mitocondrii,reticul
sarcoplasmic sla- dezvoltat,putine capilare, nivele scazute de
mio'lo-ina,rezerve de 'lico'en crescute,ATP7aza miozinica rapida,
viteza de contractie crescuta.
<licolitice rapide o-osesc rapid, au de &7( ori mai multa orta,eort
intens de scurta durata cu perioade prelun'ite de recuperare.
brele musculare roii (de tip I) con'in sarcoplasm cu mult miolo0in$
rezerve de
licoen i triliceride n cantitate mai mare dec&t la +0rele al0e. au un
numr mai mare de CG mitocondrii i un 0oat con'inut de enzime respiratorii.
<le au un meta0olism predominant anaero0$
se contract lent cu mare putere i o0osesc reu. acest tip de +0re predomin
n muc,ii tonici.
- brele albe +de tip IIB au sarcoplasma n cantitate mai mic cu miolo0in
pu'in. au un
meta0olism predominant anaero0$ 0azat pe licoliz i producerea de acid
lactic. Meta0olismul
+0relor al0e este de dou trei ori mai activ dec&t al +0relor roii i asiur
eli0erarea prompt a
eneriei de contrac'ie. <le se caracterizeaz prin contrac'ii rapide$ dar
o0osesc uor. #i0rele al0e
G8
predomin n muc,ii -e%ori$ cu contrac'ie rapid.
%8(.contractia izotona3izometrica
5ontrac'ia izometric modi+c tensiunea muc,iului$ dar lunimea rm&ne
constant. 5aracterizeaz musculatura postural. 1u produce lucru mecanic$ ci
cldur.
5ontrac'ia izotonic este aceea n care tensiunea rm&ne constant$ dar variaz
lunime. <ste caracteristic maBorit'ii muc,ilor sc,eletici. 6ealizeaz lucru
mecanic i produce micare
:n activitatea o0inuit$ muc,iul trece prin (aze de contrac'ie izometric i
izotonic$ ini'ierea oricrei contrac'ii +ind$ de o0icei$ izometric.
5ontractia izotonica: are loc atunci cand dupa cresterea tensiunii +0relor
musculare urmeaza apropierea capetelor de insertie. <ste cel mai des ntalnita n
activitatea sportiva.
5ontractia izometrica are loc atunci cand capetele de insertie ale musc,ilor si
pastreaza aceeasi distanta c,iar si dupa cresterea tensiunii +0relor musculare.
contracie izometric! atunci c&nd lunimea muc,iului rm&ne nesc,im0at
dar
tensiunea din interiorul muc,iului crete (oarte mult. :n timpul contrac'iei
izometrice$ muc,iul nu
presteaz lucru mecanic$ toat eneria c,imic se pierde su0 (orm de
cldur. Mn e%emplu de
contrac'ie izometric este aceea a muc,ilor ce(ei$ care sus'in capul n pozi'ie
normal n sta'iune
0iped.
- contracie izotonic n care lunimea muc,iului variaz (+0rele se
scurteaz) iar
tensiunea rm&ne constant.
%8).activitatea re/e1a medulara
%8*.usul neuromuscular
ST6M5TM6A:
2-8H +0re musculare intra(usale care prezint:
o por'iune central - 0oat n nuclei i termina'ii nervoase senzitive
dou por'iuni peri(erice - contractile
- cu termina'ii motorii proprii de tip
+0rele musculare e%tra(usale dispuse Nn paralelO cu +0rele musculare
intra(usale
capsul conBunctiv
0?ASI8I0ARE
$. 8I6RE I=TRA8LSA?E AdupG modul de dispunere a nucleilor Un
porF. centralGB2
fbre cu sac nuclear 7 cu nucleii a'lomeraFi Un
zona centralG
fbre cu lan nuclear 7 cu nucleii dispuIi Untr7un
sin'ur rHnd
%. 8I6RE?E SE=5ITI"E 7 terminaFii dendritice mielinizate cu
ori'inea Un 'an'lionul spinal
Fibrele senzitive primare 7 de tip Ia:
predominG la nivelul f-relor cu sac nuclear
au o dispoziFie WspiralatGX
conduc e1citaFia cu vitezG mare, de 8!7$%!
m3sec
asi'urG rGspunsul de tip dinamic 7 descarcG
impulsuri cu o recvenFG direct proporFionalG cu
G2
variaFia de lun'ime a porFiunii centrale pe
unitatea de timp
Fibrele senzitive secundare 7 de tip II:
predominG la nivelul f-relor cu lanF nuclear
au o dispoziFie Un W-uc+etX
conduce e1citaFia cu vitezG mai micG, de &!7,!
m3sec
asi'urG rGspunsul de tip static 7 descarcG
impulsuri cu o recvenFG dire
&. 8I6RE?E #OTORII 7 sunt a1onii motoneuronilor spinali de
tip , determinG contracFia porFiunilor perierice al usului Ii
creIte sensi-ilitatea f-relor senzitive la Untinderea porFiunii
centrale2
Fibre motorii dinamice
se distri-uie e1clusiv la nivelul f-relor cu sac
nuclear
asi'urG componenta azicG AdinamicGB a
re/e1ului miotatic
motoneuronii de ori'ine se a/G su- controlul
centrilor nervoIi motori supramedulari
Fibre motorii statice
se distri-uie pe am-ele tipuri de f-re intrausale
asi'urG componenta tonicG AstaticGB a re/e1ului
miotatic
motoneuronii de ori'ine se a/G su- control
strict medular
%8,.or'anul tendinos <ol'i
este un receptor pasiv, dispus WUn serieX aFG de f-rele
tendinoase
are distri-uite f-re senzitive de tip I-, cu ori'inea Un 'an'lionul
spinal
uncFioneazG ca du-lu receptor mecanic 7 de Untindere Ii de
tensiune2 descarcG impulsuri la Untinderea tendonului ApercuFia
tendonuluiB Ii la punea Un tensiune a tendonului ca urmare a
contracFiei musculare
%88.o-oseala musculara
5ontrac'iile puternice de lun durat ale muc,iului conduc la instalarea
(enomenului
+zioloic denumit o0oseal muscular. >0oseala muscular este o stare
+zioloic reversi0il ce se mani(est printr-o diminuare a activit'ilor
muc,iului dar care dispare prin repaus. Simptomele
o0oselii au (ost mpr'ite n o0iective i su0iective.
#emnele subiective sunt :
- senza'ia de reutate n micare.
dureri musculare.
- epuizare (moleeal).
#emnele obiective sunt:
- o0oseala diminueaz e%cita0ilitatea$ puterea i durata n timp a contrac'iei
musculare$
prin scderea numrului de unit'i motorii antrenate n actul motor.
- amplitudinea +ecrei contrac'ii este diminuat de o0oseal$ at&t prin
scderea numrului
GA
de +0re musculare stimulate c&t i prin reducerea capacit'ii de scurtare a
+ecrei +0re.
- diminuarea preciziei micrilor i apari'ia unor tremurturi$ consecin'a
o0oselii nervoase.
- creterea tonusului muscular n repaus.
- semnul caracteristic al o0oselii musculare este caracterul reversi0il al
acestor modi+cri.
Mecanismele o0oselii musculare rm&n (oarte controversate. se invoc (actori
locali$
0ioc,imici$ circulatori i (actori e%tramusculari relatori. "recizarea pro0lemei
o0oselii este de mari
importan' pentru +zioloia e%perimental a e(ortului i muncii$ pentru sport$
pentru medicina
clinic de e%plorri (unc'ionale i de recuperare.
- > prim teorie acord (enomenelor c,imice (epuizarea rezervelor energetice
i creterea
deeurilor acide) rolul principal n diminuarea randamentului muc,iului
o0osit$ consider&nd c
unele modi+cri electrice din muc,i i nerv sunt secundare.
Teoria nervoas (sus'inut de YrandBean) consider c la nivelul scoar'ei
cere0rale
o0oseala muscular este resim'it su0 (orma unei senza'ii speci+ce care are
drept consecin'
diminuarea numrului i (recven'ei descrcrilor de neuroni motori.
YrandBean accept diviziunea
clasic n: oboseala muscular ce se traduce prin ncetinirea i reducerea
contrac'iei musculare$
av&nd la 0az epuizarea rezervelor eneretice. oboseala mintal cu o 0oat
simptomatoloie
su0iectiv cu un mecanism predominant neuroendocrin.
9ucrurile se complic dac 'inem seama c muc,iul o0osit prin contrac'ii
voluntare
rspunde totui prin contrac'ii la stimularea electric direct a nervului su
motor. :n plus muc,iul
o0osit prin e%cita'ii directe sau indirecte i reia activitatea mai repede n
mediul alcalin prin
administrare de adrenalin sau e%citarea nervilor simpatici
Se mani(est prin diminuarea capacit'ii de travaliu muscular. Se datoreaz
scderii randamentului eneretic$ acumulrii de acid lactic$ lipsei de >2$ epuizrii
su0stan'elor macroerice i a mediatorilor c,imici la nivelul plcilor motorii.
%89.ototransductia
%9!.oc+iul emetrop si viciile de reractie
Emetropia reprezinta 7 #ocalizarea pe retina a imaini o0iectelor a-ate la
distanta cand
musc,iul ciliar este complet rela%at
>c,iul prezinta un raport corect intre puterea de re(ractie a mediilor si lunimea
a%ului
/n mod normal lunimea a%ului este de apro%imativ 28-2A mm."uterea de
re(ractie a corneei este de apro%imativ AG-CC dioptrii
si a cristalinului de 8G-2A dioptrii
%9$.vederea cromatica
Vederea cromatica
GC
r 5elulele cu conuri sunt responsa0ile pentru vederea colorata.
r 5elulele cu conuri contin (otopimenti asemanatori rodopsinei care raspund
optim
la o anumita valoare a lunimii de unda a luminii:
a) al0astru (a0sortie ma%ima C8Gnm)
0) verde (a0sortie ma%ima ?AHnm)
c) rosu (a0sortie ma%ima ?3Hnm)
<%ista doua teorii care incearca sa e%plice vederea colorata:
a) Teoria tricromatica.
0) Teoria =erin sau teoria culorilor oponente al0enal0astru.
Toti cei trei pimenti sunt necesari pentru vederea cromatica corecta.
<ste necesar minim doi pimanti pentru vederea cromatica
Vederea tricromata se intalneste la GHR din populatie prin utilizarea n proportii
apropiate de
rosu$ verde$ al0astru.
Discromatopsia este data de a0senta unui dintre pimenti$ totusi individul vede
aproape toate culorile utilizand doar doi pimenti.
Vederea monocromata implica a0senta conurilor si este (oarte rara.
Teoria tricromatica
r Se 0azeaza pe posi0ilitatea perceptie tuturor nuantelor culorilor prin
amestecarea in di(erite com0inatii ale celor trei culori (undamentale
r 5el putin doua tipuri de celule cu conuri tre0uie e%citate
r /ntensitate e%citatie +ecarui piment este comparata de catre
5reier
Teoria culorilor complementare
al0en-al0astru
r "ropune e%istenta unui proces neuroloic care considera culorile a + opuse
r 5uloarea al0astra este opusa culorii al0ene (6-V)
r 5uloarea rosie este opusa culorii verzi
r 5ele doua teorii constitue 0aza vederii colorate
%9%.acuitatea vizuala
6eprezinta capacitatea oc,iului de a discrimina 2 puncte lumininoase distincte
<ste ma%ima la nivelul (oveei +ind de 2? sec de arc de cerc si scade de 8H ori la
peri(erie
Se testeaza cu aButorul optotipului de departe
/n mod normal o persoana sanatoasa poate citi primul rand al optotipului de la
?Hm$ al doilea de la CH$ AH$2H iar ultimul de la ? m. 5itirea se (ace de la ? m unde
un su0iect sanatos poate citi ultimul rand. <%primarea se (ace printr un raport
dintre distanta la care se (ace citirea si distanta la care ar tre0ui sa poata + citit
ultimul rand.
%9&.adaptarea la intuneric si lumina
Adaptarea la intuneric
5are este e%plicatia pentru durata mare a adaptarii la intuneric.
6ecompunerea lenta a retinalului
P in intuneric retinalul este redus la (orma //-cis retinal
P se recompune rodopsina la nivelul discurilor
P nivelul de YM"c este re(acut
P canalele 1a24 se redesc,id
Pcelulele se depolarizeaza
P transmisia este reluata
%9(.acomodatia si re/e1ul pupilar
5apacitatea cristalinului de a-si modi+ca puterea de re(ractie pentru a asiura o
imaine clara atat la
G?
distanta cat si aproape.
5ristalinul are o putere de re(ractie la tanar intre 2H-AC dioptrii$ la adultul de CH
ani intre 2H-2A dioptrii si la varstnic de 2H ! 28 dioptrii
"entru realizarea acomodatie
participa: 5ristalinul$9iamentul suspensor (2H de
zonule@liamente)
V<D<6< 9A D/STA1TA
-Musc,iul ciliar (componenta circulara este cea mai imp.)
- 5ontrolul: parasimpatic (cel mai important)$ simpatic (importanta secundara)
%9).campul vizual
7reprezinta aria care poate f cuprinsa de oc+i cand acesta priveste f1
inainte.Evident, campul vizual este limitat superior de arcada
sprancenoasa, medial de piramida nazala iar inerior de pometii
o-ra.ilor. ?ateral campul vizual este desc+is, el putand trece de $$!
'rade. @atorita dispunerii dierite a celulelor cu conuri pe supraata
retinei, campul vizual este dierit pentru cele & culori undamentale.
0ampul vizual poate dieri fziolo'ic de la individ la individ, le'at de
particularitatile aciesului. Tul-urarile ma.ore sunt reprezentate de
scotoame, si in'ustari ale campului vizual.
Scotoamele reprezinta deecte de camp vizual, zone oar-e in care
su-iectul nu percepe lumina. E1ista un scotom fziolo'ic in campul vizual
dat de pata oar-a a retinei.
5orte%ul vizual
-5orte%ul vizual primar
- >cupa aria +surii calcarine si se e%tinde catre polul occipital
- Semnalele din macula au proiectie in apropierea polului occipital
-Semnalele din retina peri(erica se proiecteaza in cercuri concentrice anterior
de pol
- Structura: coloane neuronale cu diametru de AH-?H nm continand +ecare peste
8HHH de neuroni$ semnalele vizuale aBunand in stratul C si de aici urca spre
straturile 8$ 2$ A curaspandire laterala pe distante scurte$ +e co0oara in
straturile ?$ 3 cu raspandire laterala pe distante mari
-"icaturile de culoare K zone a-ate intre coloanele primare$ primesc semnale
laterale si raspund speci+c la semnalele de culoare
- Arii corticale secundare
- Situate anterior superior si in(erior de corte%ul primar vizual
- Analiza si interpretarea imainilor:
/nervatia musculaturii e%trinseci a lo0ului ocular este asiurata de nv A$ C$ 3$
/ntercone%iuniile dintre cei trei nuclei ai nv A$ C$ 3 se realizeaza prin intermediul
(asciculului lonitudinal medial
Miscarile voluntare sunt asiurate de arii din lo0ul (rontal prin tracturile
(rontotectale
Miscarile involuntare de +%atie sunt controlate de arii secundare din lo0ul
occipital prin tracturile occipitotectal si occipitocolicular
De la nucleii pretectali si coliculari sup pornesc semnale prin (asciculul
lonitudinal median spre nucleii nervilor A$ C$ 3.
%9*.in+i-itia laterala
%9,.determinarea directei sunetului
Sunetul mai este receptat i pe alte ci cum ar + calea otic osoas (mastoid-
co,lee) numit i conducerea osoas ce este para+zioloic la omul normal.
5alea osoas devine +ziopatoloic atunci c&nd este 0locat complet sistemul de
transmisie (de e%emplu$ n aeneziile meato-cavotimpanice). Mai e%ist i o
posi0ilitate de conducere cartilainoas pe cartilaBul traal (asemntoare cu
conducerea osoas). Sunetele mai pot + receptate i tactil$ palpator (n special
G3
cele rave ! de e%emplu la 83 =z)$ dar i palestezic (sensi0ilitatea vi0ratorie
osoas). Aceast receptare nueste ns n mod normal transco,lear. Mrec,ea i
nervul auditiv stau la 0aza mecanismului prin care un semnal acustic din spa'iu
este trans(ormat n poten'iale neuronale de ac'iune. Mndele sonore se propa n
tot spa'iul din Burul e%tremit'ii ce(alice iar sc,im0rile de presiune din 5A<
determin vi0ra'ia MET. Aceasta declaneaz micarea
oscioarelor. Micarea scri'ei pe (ereastra oval determin sc,im0area presiunii
n -uidele din urec,ea intern$ vi0ra'ia mem0ranei 0azilare i a oranului 5orti.
Deplasarea stereocililor celulelor senzoriale este stimulul ce duce la
trans(ormarea vi0ra'iei mecanice. /mpulsurile nervoase se propa prin a%onii
nervului auditiv. di(eren'ele de intensitate sonor dintre cele 2 urec,i date de
e(ectele Ode con(uzieO si Nde um0rO pentru stimulii de (recven' nalt depind
(oarte mult de pozi'ia sursei sonore n spa'iu
distan'a dintre urec,i determin o di(eren' de timp ntre sosirea sunetelor ntr-
o urec,e si n alta (celulele din sistemul nervos sunt (oarte sensi0ile la aceste
di(eren'e mici de intensitate si timp interaural)
direc'ia sunetului din Burul capului contri0uie la localizarea sunetului
unele specii de animale au control voluntar asupra urec,ii e%terne si pot aBusta
aceste caracteristici in avantaBul lor.
Vi0ra'ia scri'ei n (ereastra oval (ca un piston) cauzeaz
6olul urec,ii medii o di(eren' de presiune ntre (ereastra ovala si cea rotund
P Mrec,ea medie serveste ca un trans(ormator$ mreste eneria sonor
transmis -uidelor co,leare
%98.determinarea intensitatii sunetului
Deoarece variatia intensitatii sunetului (respectiv puterea sunetului) perceput
de urec,ea umana variaza (oarte mult (de la 8Ht82 a@m2 la 8H3
a@m2) intensitatea se e%prima in mod o0isnuit ca loaritmare a raportului dintre
intensitatea sunetului si intensitatea sunetul de re(erinta
%99.determinarea recventei sunetului
&!!.receptorii auditivi si transductia semnalului sonor
&!$.rolurile urec+ii medii
6ol pasiv
P "oate in-uenta propaarea sunetelor in spatiu timpul n care semnalul sonor
aBune la MET
P #orma capului$ a pavilionului i a 5A< pot a(ecta intensitatea sunetului
P "avilionul i 5A< ac'ioneaz ca i camere de rezonan' acustic a(ecteaz
presiunea
aerului la nivelul MET
P 9a sunetele cu (recven' nalt 5A< ac'ioneaz ca un simplu tu0 rezonator
desc,is la
un capt
P 9unimea acestuia determin n principal caracteristicile de rezonan' (ma%im
la om la2.?S,z)
&!%.rolul canalelor semicirculare
Structurile ce stau la 0aza mecanismului vesti0ular de men'inere a ec,ili0rului
sunt reprezentate de canalele semicirculare$ utricula i sacula. 9a nivelul utriculei
i saculei oranul receptor este
oranizat su0 (orma maculelor i este receptiv la modi+cri ale
accelera'ieiliniare.
5upula:
GL
are o densitate apropiata p p de a endolim(ei
se mic su0 ac'iunea endolim(ei ce se supune leii iner'iei de-ect&nd cilii
senzoriali n direc'ia
curerii endolim(ei
&!&.rolul utriculei si saculei
Sunt cavit'i mem0ranoase
P 5on'in +ecare o arie mic senzoriala (macula) de apro%imativ 2 mm situat n
plan orizontal
pentru utricul i respectiv n plan vertical pentru sacul #iecare macul este
(ormat din
celule paroase acoperite de o mem0ran elatinoas (otolitic) care con'ine
cristale mici de
car0onat de 5a i proteine !otoli'i
P Macula utriculei detecteaz micarea capului n plan orizontal iar macula
saculei n plan vertical
Macula saculei si a utriculei: <ste (ormat din celule paroase care prezint:
?H ! LH cili numi'i stereocili
8 cil voluminos si mai lun numit Sinocil
sunt leate n partea superioar prin +lamente de atasare su0'iri
Deplasarea cililor n direc'ia Sinocilului produce desc,iderea canalelor de U si de
5a cu producerea depolarizrii celulelor proase
P Deplasarea cililor n sens opus Sinocilului determin ,iperpolarizarea celulei
P 5elulele proase descarc neurotransmi'tor care in-uen'eaz rata de
descrcare a +0relor
vesti0ulare nervoase componente a nervului vesti0uloco,lear V///: 6epaus: 8HH
imp@sec
r Depolarizare: Z8HHimp@sec
r =iperpolarizare: f 8HH imp@sec
&!(.conducerea aeriana
Audi'ia este (unc'ia urec,ii de a percepe sunetele de anumite (recven'e si
intensit'ii
Mn sunet poate + receptat pe cale otic +zioloic numit calea aerian
"raul minim al unui auz normal pentru conducerea aerian este situat
audiometric ntre H si AH dE
Sunetul mai este receptat si pe alte ci cum ar + calea otic osoas (mastoid-
co,lee) numit si conducerea osoas ce este para+zioloic la omul normal.
5alea osoas devine +ziopatoloic atunci c&nd este 0locat complet sistemul de
transmisie (de e%emplu$n aeneziile meato-cavotimpanice).
Mai e%ist si o posi0ilitate de conducere cartilainoas pe cartilaBul traal
(asemntoare cu conducerea osoas).
pe cartilaBul traal (asemntoare cu conducerea osoas).
Sunetele mai pot + receptate si tactil$ palpator (n special cele rave ! de
e%emplu la 83 =z)$ dar si palestezic (sensi0ilitatea vi0ratorie osoas). Aceast
receptare nu este ns n mod normal transco,lear
&!).corpusculul Pacini2structura,caracteristici fziolo'ice
localiza'i n 'esutul su0cutanat i dermul pro(und
Pprezen'i n pielea la0r i cea proas
Pnumeroi n ,ipodermul palmelor i plantelor
Pvoluminoi - lunime apro% 2 mm.
P(orm eliptic
Alctui'i din:
capsul (ormat din 2H-3H lamele concentrice de 'esut conBunctiv ntre
GD
care e%ist un -uid vscosKZ aspect asemntor (oi'elor de ceap
+0r nervoas care se termin printr-o um-tur n contact cu lamelele
centrale. captul +0rei e nemielinizat$ dar cu pu'in nainte de a prsi
corpusculul$ +0ra devine mielinizat. primul nod 6anvier e situat de asemenea n
interiorul corpusculului.
5orpusculii "acini se adapteaz (oarte rapid$ n c&teva sutimi de secund
Stimul aplicat lent$ sau care ac'ioneaz continuu KZ rearanBarea lamelelelor
interne ale capsulei KZ termina'ia nervoas nu mai este de(ormat.
Doar de(ormrile rapide sunt transmise e+cient ctre reiunea central a
capsulei.
De +ecare dat c&nd stimulul este aplicat i apoi ndeprtat corpusculul "acini
descarc poten'iale de ac'iune$ (recven'a de descrcare +ind propor'ional cu
(recven'a stimulului vi0rator
5orpusculii "acini - importan'i n perceperea senza'iei Vi0ratorii
5orpusculii "acini
Ppercep vi0ra'ii cu (recven'e ntre AH i DHH cicli@sec
Psunt e%trem de sensi0ili ! rspund la vi0ra'ii cu amplitudine de su0 8H nm la
supra(a'a pielii KZ implica'i n detectarea caracteristicilor unor supra(e'e prin
intermediul unor unelte 'inute n m&n
&!*.sensi-ilitatea tactila discriminativa
distan'a minim ce tre0uie s separe doi stimuli pentru ca acetia s +e
percepu'i separat.
Manitudinea sa variaz de la o reiune la alta a corpului:
-v&r(ul lim0ii: 8$8 mm
-deete ! (a'a palmar: 2$2mm
-0uze: C$? mm
-v&r(ul nasului: 3$D mm
-(a'a dorsal a m&inii: 83 mm
-torace posterior$ coaps: 3? mm
Doi (actori importan'i:
rmrimea c&mpului receptor
rdensitatea receptorilor cutana'i
&!,.transmiterea senzatiilor tactile vi-ratorii si de presiune prin f-rele
nervilor perierici
6eceptorii senzoriali specializa'i ce permit localizarea precis pe teument$
rada'iile (oarte +ne de intensitate sau sc,im0rile rapide de intensitate a
stimulului )+0re roase$ mielinizate de tip A 0eta$
udiametru: 3-82 bm
uvitez de transmitere: AH-LH m@s.
PTermina'iile nervoase li0ere ce transmit senza'ii mai 0rute$ precum presiunea
rosier$ atinerea sla0 localizat )+0re su0'iri mielinizate tip A delta
udiametru: 8-? bm
uvitez de transmitere: ?-AHm@s.
)+0re nemielinizate de tip 5
udiametru: H$2-8$? bm
uvitez de transmitere: v2 m@s.
&!8.receptorii durerii
urealizeaz transduc'ia n impulsuri nervoase a stimulilor nocivi ce ac'ioneaz
asupra 'esuturilor
usunt termina'ii nervoase li0ere i sunt distri0ui'i n aproape toate 'esuturile
oranismului
uau densitate mare la nivel cutanat
P1ociceptorii mecanici
utermina'ii nervoase ale unor +0re milelinizate su0'iri tip A delta
GG
ustimul: presiunea$ n'eptura$ vi0ra'ia$ distensia$ trac'iunea
P1ociceptorii termici
utermina'ii nervoase ale unor +0re mielinizate su0'iri de tip A delta i +0re
nemielinizate de tip 5
ustimul: temperaturi e%treme
P1ociceptorii c,imici
utermina'ii nervoase ale unor +0re su0'iri$ nemielinizate de tip 5
urspund la varia'i aen'i precum 0radiSinina$ ,istamina$ serotonina$
acetilcolina$ enzimele proteolitice$ AT"$ ionii =4$ ionii U4$ precum i numeroase
su0stan'e iritante din mediul e%tern
P1ociceptorii polimodali
utermina'ii nervoase li0ere ale unor +0re amielinice de tip 5
usensi0ili at&t la stimuli mecanici$ termici c&t i c,imici.
PEradiSinina
activeaz direct nociceptorii - receptor speci+c E2
P=istamina
produce o durere de durat redus - receptor speci+c =8.
P<nzimele proteolitice produc durere mediat de receptorul "A6-2 (proteinase
activated
receptor 2)
PAcidoza tisular determin durere mediat de canalele de tip AS/5
P5apsaicina (inredient picant con'inut n ardeii iu'i) determin durere prin
ac'iune direct pe receptorul T6"V8.
receptori pentru durere - stimula'i numai de
temperaturile e%treme
Densitatea termoreceptorilor la nivelul pielii:
Pcea mai mare densitate de receptori termici$ at&t pentru cald c&t i pentru rece$
este la nivelul (e'ei
Pe%ist de A-8H ori mai mul'i receptori pentru rece dec&t pentru cald.
Pdensitatea punctelor sensi0ile la rece:
u0uze:8?-2H pe cm2
udeete: A-? pe cm2
utrunc,i: 8 pe cm2.
receptorii pentru rece: rami+ca'iile unor +0re su0'iri mielinizate
Preceptorii pentru cald: n cea mai mare parte rami+ca'ii ale unor +0re su0'iri
amielinice i ntr-o mic msur rami+ca'ii ale unor +0re mielinizate su0'iri.
la nivel cutanat - canale ionice sensi0ile la temperatur$ apar'in&nd super(amiliei
T6" (Transient 6eceptor "otential)
ce rspund la di(erite prauri de temperatur (T6"V8$T6"V2$ T6"VA$ T6"VC$
T6"MD$ T6"A8).
T6"V8
rcanal cationic neselectiv activat de temperaturi ZCAo5
rpoate + activat de capsaicin$ inredientul picant din ardeii
iu'i (aplicat pe piele determin senza'ie de arsur)
T6"V2
rrspunde la temperaturi mai ridicate (Z?2o5)
rnu este activat de capsaicin
T6"VA
rcanal activat de temperaturi de apro%imativ AA o5.
rcam(orul - activator speci+c al T6"VA (aplicarea de cam(orpe piele determin o
senza'ie de cldur)
rpra de temperatur Z2L-A?o5
T6"MD
rcanal implicat n senza'ia de rece
8HH
reste numit i 5M6-8 (cold and ment,ol receptor 8).
ractivat la o temperatur f 2?-2Do5
rstimulat de mentol (aplicarea de mentol pe teument
determin o senza'ia rcoroas)
T6"A8
reste activat la temperaturi mai reduse (f8Lo5)
rparticip la perceperea senza'iei de n,e'at.
ractivat i de diverse su0stan'e picante din mutar$
scor'ioar etc.
Temperaturi (oarte sczute - stimulate doar +0rele pentru durere-rece
8H 8? o5 impulsurile de durere rece diminu ncep s +e stimula'i receptorii
pentru rece
? 8H 8? 2H 2? AH A? CH C? ?H ?? 3H
Temperatura (o5)
8H-o5$ durere-diminu$ rece$
AH o5 ncep s +e stimula'i receptorii pentru cald$ (recven'a de descrcare a
impulsurilor
diminu&nd spre ?H o5.
C? o5 ncep s +e stimulate +0rele de durere-cald. apare o stimulare
parado%al a +0relor
pentru rece
r6eceptorii pentru cald transmit impulsurile n principal prin +0re nervoase
nemielinizate tip 5$ cu
viteze de transmitere de H$C-2 m@s.
r6eceptorii pentru rece transmit impulsurile prin +0re nervoase mielinizate su0tiri
tip A delta$ cu viteze de apro%imativ 2H m@s.
&!9.caracteristicile durerii rapide si ale durerii lente
rDurerea rapid
Presim'it la H$8 secunde de la aplicarea stimulului. are caracter de n'eptur
Pdeterminat n special de stimuli mecanici sau termici
Ptransmis prin +0re tip A delta cu viteze de 3-AH m@s
Pse asociaz cu re-e% de retraere$ rspuns simpatic$ creterea presiunii arteriale
i mo0ilizarea rezervelor eneretice ale oranismului
rDurerea lent
Pncepe dup mai mult de o secund$ se intensi+c lent$ este surd i are
caracter de arsur
Ppoate + determinat de stimuli mecanici$ termici sau c,imici
Ptransmis prin +0re de tip 5 cu viteze de H$?-2 m@s.
Pse asociaz cu rea'$ transpira'ii pro(uze$ scderea presiunii arteriale i
reducerea eneralizat a tonusului muscular
&$!.caracteristici ale transmiterii semnalului in sistemul lemniscal
dorsal si sistemul anterolateral
1euronii primari ai cii transmiterii sensi0ilit'ii cutanate
- anlionii rdcinilor dorsale sau anlionii ec,ivalen'i ai nervilor cranieni.
#i0rele a(erente primare ptrund la nivelul mduvei spinrii pe calea rdcinilor
posterioare a
nervilor spinali.
De la punctul de intrare n mduv$ impulsurile senzitive sunt transmise spre
creier pe una din urmtoarele ci senzitive: sistemul lemniscal dorsal i sistemul
anterolateral.
Sistemul lemniscal dorsal
8H8
+0re nervoase mielinizate roase
vitez de conducere mare - AH-88Hm@s
rad ridicat de orientare spa'ial a +0relor nervoase n (unc'ie de oriinea lor
pe supra(a'a corpului (reiunile in(erioare - medial$ reiunile superioare - lateral).
transmiterea senza'iilor cu o rada'ie a intensit'ii (oarte precis$ permi'&nd
p&n la 8HH rade de intensitate.
capacitate crescut de a transmite impulsuri repetitive.
transmite in(orma'ia senzorial rapid i cu +delitate mare temporo-spa'ial
transmite senza'iile tactile sau presionale ce necesit un rad mare de
localizare a stimulului sau discriminarea unor rada'ii +ne de intensitate$
senza'iile vi0ratorii$ micarea unui o0iect (a' de piele$ senza'ii de pozi'ie
Sistemul anterolateral
+0re mielinizate su0'iri
vitez de conducere de D-CH m@s$
rad mult mai mic de orientare spa'ial a +0relor
transmiterea senza'iilor cu o rada'ie a intensit'ii mai pu'in precis$ permi'&nd
recunoaterea a doar 8H - 2H de rade de intensitate
capacitate redus de a transmite impulsuri repetitive
transmite in(orma'ia a crei transmitere nu necesit nici rapiditate nici mare
+delitate spa'ial
transmite un spectru mai lar de modalit'i senzoriale: senza'ii tactile i
presionale rosiere$ senza'ii termice$durere$ prurit$ senza'ii se%uale.
5onducerea prin sistemul lemniscal dorsal
#i0rele senzoriale KZ r. medial i r. lateral.
r6amura lateral - su0st. cenuie medular
)cone%iuni la nivelul cornului posterior ! pot modi+ca inputul altor sisteme
senzoriale cutanate (e%: durerea)
)sinapse cu neuroni din por'iunile intermediar i anterioar ale su0stan'ei
cenuii medulare
(declanaz re-e%e medulare locale sau dau natere tracturilor spinocere0eloase)
r6amura medial - cordonul posterior )sinaps n 0ul0 (nucleii cordoanelor
dorsale - nucleii cuneat i racilis).
Deutoneuronii se ncruciaz imediat - lemniscurile mediale - talamus (comple%
ventro0azal) neuron ter'iar - aria senzorial somatic / (irusul postcentral)$ aria
senzorial somatic // (por'iunea lateral i in(erioar a +ecrui lo0 parietal)
5onducerea n sistemul anterolateral
#i0rele KZ sinapse n coarnele posterioare ! ncruciare prin comisura anterioar
a mduvei - traiect ascendent prin cordoanele anterolaterale de partea opus.
rTractul spinotalamic anterior - sensi0ilitate tactil rosier
rTractul spinotalamic lateral - durerea i temperatura
r/mpulsurile tactile i pro0a0il impulsurile termice ! n principal ctre talamus
(comple%ul ventro0azal) KZ corte%ul somatosenzorial
rDurerea rapid - ctre comple%ul ventro0azal talamic KZ corte%ul senzorial
somatic i arii 0azale ale
creierului
rDurerea lent - ctre trunc,iului cere0ral (nuclei reticulari$ aria tectal
mezence(alic i su0stan'a
cenuie periapeductal) KZ nucleii intralaminari
talamici i anumite reiuni din ,ipotalamus.
"erceperea durerii - n principal (unc'ia centrilor in(eriori
/nterpretarea calit'ii durerii. - corte%ul
&$$.receptorii si transductia senzatiei 'ustative
&$%.receptorii si transductia senzatei olactive
&$&.calea autonoma simpatica
8H2
Dominan'a activit'ii simpatice este cauzata de stresul emo'ional
sau +zic
! uren'e$ nriBorare$ (rustrare$ e%cita'ie$ e%erci'iu
P 6eac'ii de alarm K rspuns (u sau lupt
! Dilata'ie pupilar
! <(ect cronotrop 4$ inotrop 4 w vasopresor 4
! Diminuarea -u%ului sanvin n orane neesen'iale
5reterea per(uziei sanvine n muc,iul sc,eletic i cardiac
! ! Eron,odilata'ie w creterea ratei respiratorii
! 5reterea licemiei w AY circulan'i
P <(ect de durat datorat persisten'ei 1A in (anta sinaptic i eli0errii
1A din l. MS6 #aciliteaz procesele de asimilare i recuperare
P Activeaz mecanismele de conservare i reenerare a eneriei n repaus
P #izioloic domin peste tonusul simpatic
P 5ontroleaz procese de tip saliva'ie$ secretie lacrimala$ urinare$ diestie
de(eca'ie diestie$
P A procese de scdere ! (recven' cardiaca$ diametrul cilor aeriene i diametrul
pupilar
P Team "arado%al ! n situa'ii (r scpare@disperate
! Activare masiv a componentei parasimpatice
! "ierderea controlului s+ncterian (mic'iune@de(eca'ie involuntar)
&$(.calea autonoma parasimpatica
&$).receptorii adrener'ici
1euronii adrenerici eli0ereaz noradrenalina (1A@1<) 1MMA/ din neuronii
postanlionari simpatici
P Ac'iune: e%cit sau in,i0it oranele 'int n (unc'ie de receptori
P 6eceptori:
! Al(a2 produc in,i0i'ie
! Al(a8$ Eeta8 i Eeta2 produc e%cita'ie
! EetaA ('esut adipos 0run) cresc termoeneza
P 1A persist la nivel sinaptic pana c&nd este inactivat enzimatic de
monoamine-o%idaz (MA>) sau catecol->-metil-trans(eraz (5>MT)
&$*.receptorii coliner'ici
6eceptorii nicotinici musculari em0rionari
- pe toat supr(a'a +0rei musculare neinervat sau denervat
/.Distri0u'ia receptorilor colinerici
- au o rat mare a turnoverlui
6eceptorii nicotinici musculari Bonc'ionali
- la nivelul Bonc'iunii neuromusculare
- au o rat mic a turnoverlui
6eceptorii nicotinici neuronali
- simpatici i parasimpatici$se a- n creasta neural$au propriet'i identice
- postsinaptici n anlionii simpatici i parasimpatici i medulosuprarenal
6eceptorii muscarinici
- la nivelul neuronilor postanlionari parasimpatici
- inerveaz m.neted visceral$miocard i glande secretorii
-n centrii nervoi superiori la nivelul interneuronilor-corpul striat- controlul motor extrapiramidal i
memorie
-n centrii nervoi superiori la nivelul cel. Renshaw n centrii nervoi superiori la nivelulneuronilor de
proiecie din nc.bazali
&$,.medierea autonoma la nivelul eectorilor
&$8.medulosuprarenala
Anatomic i (unc'ional medulosuprarenala este un anlion simpatic iant$ al
crui celule
8HA
postanlionare nu au preluniri i secret (ca i neuronii simpatici
postanlionari) catecolaminele
(adrenalina DH R i noradrenaliana 2H R) pe care le vars n s&ne. ea se
dezvolt din ectodermul
crestelor anlionare.
Ac'iunea acestor ,ormoni este identic cu e(ectele e%cita'iei sistemului
nervos simpatic. de
(apt termina'iile simpatice eli0ereaz aceleai catecolamine$ dar n propor'ie
invers. "rincipalele
ac'iuni ale acestor ,ormoni (care sunt i mediatori c,imici ai sistemului
nervos simpatic) sunt:
)noradrenalina determin contrac'ia musculaturii netede din peretele vaselor
(vasoconstricie)$
rele%area musculaturii netede$ n paralel cu contrac'ia s+ncterelor de la
nivelul tu0ului diestiv$ ca
i de la nivelul vezicii urinare.
)adrenalina are e(ect di(eren'iat asupra musculaturii netede a vaselor din
di(erite teritorii. ea
produce vasoconstricie$ la nivelul teumentului i a oranelor interne$ n
paralel cu vasodilataie$
la nivelul vaselor din muc,ii stria'i$ ca i la nivelul coronarelor. Ac'ion&nd
asupra inimii$
adrenalina are drept e(ect o cretere a (or'ei de contrac'ie a muc,iului
cardiac i o creterea
(recven'ei de contrac'ie$ ceea ce are drept consecin' creterea de0itului
cardiac.
Adrenalina are e(ecte predominant meta0olice i eneretice$ ac'ion&nd
asupra
meta0olismului intermediar prin stimularea licolizei i lipolizei$ ce produc
licoenoliz i
,iperlicemie$ mo0iliz&nd rsimile din depozitele adipoase i cata0olismul
acizilor rai li0eri din
s&ne. Ac'iunea adrenalinei se mani(est at&t prin creterea tonusului
centrilor nervoi veetativi
din trunc,iul cere0ral$ c&t i prin creterea e%cita0ilit'ii neuronilor corticali.
&$9.re'larea secretiei medulosuprarenalei
6elarea secre'iei medulosuprarenalei se (ace prin mecanisme neuroumorale.
6olul de
stimulator l are sistemul nervos simpatic i licemia. scderea licemiei
stimuleaz secre'ia de
catecolamine$ iar creterea licemiei o diminu. Stress-ul i suprasolicitrile
stimuleaz secre'ia
medulosuprarenei. :n somn i n condi'ii 0azale$ secre'ia este sczut. :n e(ort
+zic$ la (ri$ n
,ipotensiune$ emo'ii$ se produce o secre'ie mrit de catecolamin$ n care
propor'ia celor 2
,ormoni se poate sc,im0a: n stress-uri cu care individul este o0inuit crete
noradrenalina$ iar n
stress-uri neo0inuite crete adrenalina.
Scderea concentra'iei sanuine a ,ormonilor corticoizi i creterea concentra'iei
de noradrenalin secretat de medulosuprarenala stimuleaz adeno,ipo+za n
sensul sporirii secre'iei de ,ormon corticotrop A5T= (adrenocorticotrop). Acesta
8HC
stimuleaz corticosuprarenala n sensul secre'iei de ,ormoni corticoizi. 5reterea
concentra'iei sanuine a ,ormonilor corticoizi in,i0 secre'ia de A5T= din partea
adeno,ipo+zei$ iar scderea concentra'iei de A5T= are ca e(ect reducerea
secre'iei de ,ormoni corticoizi din partea corticosuprarenalei
6elarea secre'iilor medulosuprarenalei se realizeaz independent de ,ipo+z$ pe
cale neuroumoral i nervoas (v. meta0olismul lucidelor). "e cale nervoas$
prin intermediul scoar'ei$ are loc adaptarea secre'iilor medulosuprarenalei i de
aici activitatea visceral la solicitrile mediului e%tern.
&%!.modifcari autonome in somn
P =ipotonie muscular eneralizat (cu e%cep'ia sistemelor cardiovascular$
respirator i oculo-motor).
P Micri oculare rapide n special dreapta ! st&na.
P Activarea n propor'ie de LH ! DHR a masei neuronale cere0rale$ crete
meta0olismul$
vasculariza'ia$ volumul cere0ral$ temperatura cu 8o5
P <%cita'ie a oranelor se%uale.
&%$.modifcari fziolo'ice in raspunsul la stres
P 5&nd e%ist un volum mare de in(orma'ii de asimilat$ nceputul vie'ii
pro(esionale.
P :nceputul vie'ii de rela'ie (curtare$ cuplu$ etc.).
P :n sarcin.
P :n stresul acut.
P :n situa'ii de izolare.
8H?
S-ar putea să vă placă și
- Sistemul CirculatorDocument17 paginiSistemul Circulatorgogulatorul90% (10)
- Legea Frank StarlingDocument9 paginiLegea Frank StarlingIonuț CozmaÎncă nu există evaluări
- Encefalopatie HTDocument30 paginiEncefalopatie HTMihaela Ionela OnofreiÎncă nu există evaluări
- Carte Ro 03 04 2009Document524 paginiCarte Ro 03 04 2009Merry LeeÎncă nu există evaluări
- Lucrare Intreaga Socul CardiogenDocument77 paginiLucrare Intreaga Socul CardiogenAna Maria MaximÎncă nu există evaluări
- Curs CardiologieDocument21 paginiCurs CardiologieVioleta DragomirÎncă nu există evaluări
- Circulație - Activitatea CardiacăDocument8 paginiCirculație - Activitatea CardiacăManuela ConcioiuÎncă nu există evaluări
- Reglarea Neuro-Umorala A Activitatii Inimii. Sindromul Shy-DragerDocument11 paginiReglarea Neuro-Umorala A Activitatii Inimii. Sindromul Shy-Dragerstefan2415Încă nu există evaluări
- Partea A II - A - Anatomia Si Fiziologia Sistemului Circulator.Document6 paginiPartea A II - A - Anatomia Si Fiziologia Sistemului Circulator.Mitu b.i.Încă nu există evaluări
- Boala HipertensivaDocument42 paginiBoala HipertensivahoidragÎncă nu există evaluări
- Tema 4.fiziologia Sistemului Cardiovascular 2Document101 paginiTema 4.fiziologia Sistemului Cardiovascular 2Ionela ZubcoÎncă nu există evaluări
- Medicatia Aparatului Cardiovascular AntianginoaseDocument49 paginiMedicatia Aparatului Cardiovascular AntianginoaseDaniGFÎncă nu există evaluări
- Aritmii ElectrofiziologieDocument30 paginiAritmii ElectrofiziologiePatricia PatzÎncă nu există evaluări
- Splina: Organ Abdominal Ce Apartine Sistemului Circulator SanguinDocument5 paginiSplina: Organ Abdominal Ce Apartine Sistemului Circulator SanguinLaura PadureanÎncă nu există evaluări
- Sistemul Cardiovascular Și AnesteziaDocument42 paginiSistemul Cardiovascular Și AnesteziaKim TaehyungÎncă nu există evaluări
- CDFDocument6 paginiCDFValeria JmÎncă nu există evaluări
- Ciclu CardiacDocument8 paginiCiclu CardiacClaudiaÎncă nu există evaluări
- Ciclul CardiacDocument5 paginiCiclul CardiacGabriel HerbeiÎncă nu există evaluări
- Sistemul CirculatorDocument8 paginiSistemul CirculatorbrailajennyÎncă nu există evaluări
- Hipotensiunea ArterialaDocument62 paginiHipotensiunea Arterialageorgealink100% (1)
- Sângel E: As - Univ.Dr - Horatiu NeagosDocument19 paginiSângel E: As - Univ.Dr - Horatiu NeagosCraciun CatalinaÎncă nu există evaluări
- FIZIOLOGIA APARATULUI CARDIOVASCULARDocument13 paginiFIZIOLOGIA APARATULUI CARDIOVASCULARAdrian Gîldău100% (1)
- Medicaţia Aparatul Cardiovascular - TonicardiaceDocument55 paginiMedicaţia Aparatul Cardiovascular - TonicardiaceEdwardRazvanÎncă nu există evaluări
- Fizio LP - Revolutia Cardiaca 2Document2 paginiFizio LP - Revolutia Cardiaca 2Ana-Maria CroitoruÎncă nu există evaluări
- Sistemul CirculatorDocument10 paginiSistemul CirculatorPetrina RusuÎncă nu există evaluări
- 19 Circulatie InimaDocument5 pagini19 Circulatie InimaDana Iulia MoraruÎncă nu există evaluări
- Fiziologia InimiDocument5 paginiFiziologia InimiCiprian DicuÎncă nu există evaluări
- FiziopatDocument23 paginiFiziopatDianu MadalinÎncă nu există evaluări
- Aparatul CirculatorDocument10 paginiAparatul CirculatorMarian Danila MarianÎncă nu există evaluări
- Rezumat CirculatiaDocument11 paginiRezumat CirculatiaElena CiobanuÎncă nu există evaluări
- Anatomia Si Fiziologia Sistemului Cardio-VascularDocument37 paginiAnatomia Si Fiziologia Sistemului Cardio-VascularGabrielaÎncă nu există evaluări
- Aparatul CV Si Pato - Ogie AsociataDocument65 paginiAparatul CV Si Pato - Ogie AsociataMelania ChirtesÎncă nu există evaluări
- Fiziologie TotDocument45 paginiFiziologie TotPasvantu DanielÎncă nu există evaluări
- Cardiologie Curs 1 Anatomia InimiiDocument27 paginiCardiologie Curs 1 Anatomia InimiieliÎncă nu există evaluări
- Activitatea CardiacaDocument21 paginiActivitatea CardiacaIon CreangăÎncă nu există evaluări
- Curs 3 Reglarea Activitatii CardiaceDocument4 paginiCurs 3 Reglarea Activitatii CardiaceToader IsabelaÎncă nu există evaluări
- Masurarea Si Notarea PulsuluiDocument4 paginiMasurarea Si Notarea PulsuluigrigorescucristinaÎncă nu există evaluări
- Motivatia LucrariiDocument73 paginiMotivatia LucrariiMihaela EnacheÎncă nu există evaluări
- Ciclul CardiacDocument4 paginiCiclul CardiacAmaicei Larisa100% (1)
- LP 2 MCDocument40 paginiLP 2 MCAlexandra UngureanuÎncă nu există evaluări
- Sistemul Cardiovascular3Document7 paginiSistemul Cardiovascular3Gavrilescu AlexandraÎncă nu există evaluări
- 3 Sistemul Cardio VascularDocument27 pagini3 Sistemul Cardio Vascularlena bursucÎncă nu există evaluări
- Finalmorfologia Şi Fiziologia Cordului2022Document48 paginiFinalmorfologia Şi Fiziologia Cordului2022Ana SanduÎncă nu există evaluări
- Blocurile Atrio VentriculareDocument5 paginiBlocurile Atrio Ventricularecatalin.mÎncă nu există evaluări
- Fizio LP CardiovascularDocument85 paginiFizio LP CardiovascularAna Cristina100% (8)
- 2023 - Sistemul CirculatorDocument10 pagini2023 - Sistemul CirculatorBuftea AndreeaÎncă nu există evaluări
- Subiect eDocument9 paginiSubiect eBianca StoicaÎncă nu există evaluări
- INFORMATIIDocument39 paginiINFORMATIIIozefina AvramÎncă nu există evaluări
- Investigarea Sistemului CardiovascularDocument12 paginiInvestigarea Sistemului CardiovascularVioleta FridjoiÎncă nu există evaluări
- Sis CardiovascularDocument5 paginiSis Cardiovascularrosse_maryÎncă nu există evaluări
- Functiile InimiiDocument12 paginiFunctiile InimiiClaudiu SigheartauÎncă nu există evaluări
- Curs 2.04.2012 Sistemul Renina Angiotensina Aldosteron BradikiniaDocument3 paginiCurs 2.04.2012 Sistemul Renina Angiotensina Aldosteron BradikiniacnajesrnasiferfÎncă nu există evaluări
- DisritmiiDocument120 paginiDisritmiiАлина МаковенчукÎncă nu există evaluări
- Referat GonoreaDocument106 paginiReferat GonoreaMIHAELA STANCOIÎncă nu există evaluări
- Curs 1 2 Sfredel 2009Document40 paginiCurs 1 2 Sfredel 2009Bodeanu Suzana MihaelsÎncă nu există evaluări
- Sistemul Circulator. InimaDocument4 paginiSistemul Circulator. InimaMARINELA PETRIEÎncă nu există evaluări
- Inima CursDocument9 paginiInima Cursvanessa dinuÎncă nu există evaluări
- Microcirculația sanguină: Următoarea generație de asistență socialăDe la EverandMicrocirculația sanguină: Următoarea generație de asistență socialăÎncă nu există evaluări
- File alese din istoria cardiologiei romanestiDe la EverandFile alese din istoria cardiologiei romanestiÎncă nu există evaluări
- Kineto MedularDocument70 paginiKineto MedularMerry LeeÎncă nu există evaluări
- Kineto AvcDocument58 paginiKineto AvcMerry LeeÎncă nu există evaluări
- Kineto Cerebelos VestibularDocument42 paginiKineto Cerebelos VestibularMerry LeeÎncă nu există evaluări
- Kineto Avc 2Document69 paginiKineto Avc 2Merry LeeÎncă nu există evaluări
- Kineto ExtrapiramidalDocument44 paginiKineto ExtrapiramidalMerry LeeÎncă nu există evaluări
- Principii Terapeutice La Varstnici Mihu FlorianDocument16 paginiPrincipii Terapeutice La Varstnici Mihu FlorianMerry LeeÎncă nu există evaluări
- Ref - Supl .Locuri-Nov.20211Document4 paginiRef - Supl .Locuri-Nov.20211Merry LeeÎncă nu există evaluări
- Curriculum Stimulatoare Și Defibrilatoare Cardiace 2017Document14 paginiCurriculum Stimulatoare Și Defibrilatoare Cardiace 2017Merry LeeÎncă nu există evaluări
- Subiecte Psihiatrie-RezolvateDocument32 paginiSubiecte Psihiatrie-RezolvateMerry Lee100% (1)
- Grile FarmacologieDocument3 paginiGrile FarmacologieIrina Elena100% (1)