S-ar putea să vă placă și
- Membrane Biologice - BunDocument21 paginiMembrane Biologice - BunMarius Alexa67% (3)
- Proiect BEMADocument29 paginiProiect BEMAMarius Baluta0% (1)
- Referat Membrana CelularaDocument15 paginiReferat Membrana CelularaRazvan_GabrielÎncă nu există evaluări
- Capitol Materiale FiltranteDocument13 paginiCapitol Materiale FiltranteCristina Halbac Cotoara ZamfirÎncă nu există evaluări
- Ordinul DipteraDocument7 paginiOrdinul DipteraNeagoie DanÎncă nu există evaluări
- Recombinarea GeneticaDocument6 paginiRecombinarea Geneticamosneagu_ana_ma6397Încă nu există evaluări
- Fiziopatologia Echilibrului Fluido-CoagulantDocument5 paginiFiziopatologia Echilibrului Fluido-CoagulantSimina Elena SimionescuÎncă nu există evaluări
- Biocel - Subiecte RezolvateDocument94 paginiBiocel - Subiecte RezolvateVictorVasile95% (20)
- MICROBIOLOGIEDocument108 paginiMICROBIOLOGIEAlyn AlyÎncă nu există evaluări
- Referat GeneticaDocument6 paginiReferat GeneticaVykol AlexandruÎncă nu există evaluări
- Microbiologie Generala 2016-2Document57 paginiMicrobiologie Generala 2016-2Cristian MortuÎncă nu există evaluări
- Virusurile Ca Sisteme Microbiologice de Tip AcelularDocument17 paginiVirusurile Ca Sisteme Microbiologice de Tip AcelularEnache AlexandruÎncă nu există evaluări
- HistologieDocument39 paginiHistologieConstantin UntilaÎncă nu există evaluări
- Curs Stom Embrio1-68874Document130 paginiCurs Stom Embrio1-68874Alexandru IarovoiÎncă nu există evaluări
- Cap1-2-1 Plasmide Bacteriene STR Clasif ExtraDocument3 paginiCap1-2-1 Plasmide Bacteriene STR Clasif Extracreatza20Încă nu există evaluări
- Curs 5 Mutatii 14 OctDocument129 paginiCurs 5 Mutatii 14 OctCaraiman RalucaÎncă nu există evaluări
- Fiziologia VegetalăDocument34 paginiFiziologia VegetalăNatalia Atamaniuc100% (1)
- Helminti PlatiDocument62 paginiHelminti PlatisazunaxÎncă nu există evaluări
- ImaginiDocument18 paginiImaginiMaria RusuÎncă nu există evaluări
- 8.definiți Noțiunea de Aberație Cromozomială. Clasificarea Aberațiilor CromozomialeDocument3 pagini8.definiți Noțiunea de Aberație Cromozomială. Clasificarea Aberațiilor CromozomialeCristian TurcanÎncă nu există evaluări
- Histologie Intrebari 2019Document12 paginiHistologie Intrebari 2019Ana Cristina BitcaÎncă nu există evaluări
- Morfologia BacteriilorDocument33 paginiMorfologia BacteriilorDaniela AramaÎncă nu există evaluări
- Curs 1 MicrobiologieDocument54 paginiCurs 1 MicrobiologieFam ChirilaÎncă nu există evaluări
- Meioza - Diviziunea MeioticaDocument13 paginiMeioza - Diviziunea MeioticaIoana AgafieiÎncă nu există evaluări
- Curs Fiziologie Animala GeneralaDocument93 paginiCurs Fiziologie Animala GeneralaCristina Neg100% (1)
- Microbiologie Curs 4Document59 paginiMicrobiologie Curs 4Tocianiuc Vlad100% (1)
- Cursfito 8Document6 paginiCursfito 8BuluguTraianLucianÎncă nu există evaluări
- FiziologieDocument82 paginiFiziologieiulianspaÎncă nu există evaluări
- Virusologie GeneralăDocument55 paginiVirusologie Generalăbianca111519Încă nu există evaluări
- Penicillium AspergillusDocument2 paginiPenicillium AspergillusActurus AndreiÎncă nu există evaluări
- Holoproteide HeteroproteideDocument7 paginiHoloproteide HeteroproteideBianca IonițăÎncă nu există evaluări
- LIPOZOMIDocument11 paginiLIPOZOMIRoberta StoicaÎncă nu există evaluări
- Microbiologie Bacteriofagul T4Document6 paginiMicrobiologie Bacteriofagul T4Dan Raducu Mhl50% (2)
- Microbiologie Cs RedactatDocument24 paginiMicrobiologie Cs RedactatАлександра БантушÎncă nu există evaluări
- Genetica Microorganismelor Şi Inginerie Genetică Microbiană NoteDocument19 paginiGenetica Microorganismelor Şi Inginerie Genetică Microbiană Notecreatza20100% (4)
- FLOAREA SOARELUI Agricultura EcologicaDocument13 paginiFLOAREA SOARELUI Agricultura Ecologicasilviu_adi_1Încă nu există evaluări
- Inducerea Artificiala A PoliploidieiDocument5 paginiInducerea Artificiala A PoliploidieiFlorin AggoÎncă nu există evaluări
- Biofizica 8Document39 paginiBiofizica 8Lile GeorgeÎncă nu există evaluări
- Evaluare Corina 2Document2 paginiEvaluare Corina 2constantinÎncă nu există evaluări
- Fasciola HepaticaDocument6 paginiFasciola HepaticaRoxana AndreeaÎncă nu există evaluări
- Referat Master ParazitologieDocument28 paginiReferat Master ParazitologiearabianoudÎncă nu există evaluări
- Ciupercile FitopatogeneDocument10 paginiCiupercile FitopatogeneFlorin ArdeleanÎncă nu există evaluări
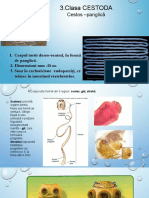
- Clasa CestodaDocument51 paginiClasa CestodaEmmaÎncă nu există evaluări
- Imunitatea Dobândită. Sistemul Imun. Celulele Imunocompetente. Cmh. Antigene. Anticorpi.Document3 paginiImunitatea Dobândită. Sistemul Imun. Celulele Imunocompetente. Cmh. Antigene. Anticorpi.anna mariaÎncă nu există evaluări
- 12 - Cresterea Plantelor IDDocument12 pagini12 - Cresterea Plantelor IDTitel BÎncă nu există evaluări
- Enzime CelulozoliticeDocument26 paginiEnzime CelulozoliticeAlexandra CabatÎncă nu există evaluări
- Planul Seminarelor La Bioet Stom 2020-2021-18381Document4 paginiPlanul Seminarelor La Bioet Stom 2020-2021-18381studentÎncă nu există evaluări
- Sapt 3 - LP Caractere Generale CerealeDocument21 paginiSapt 3 - LP Caractere Generale CerealeŞtefan Laurenţiu BătrînaÎncă nu există evaluări
- Referat MicrobiologieDocument8 paginiReferat Microbiologieliviu_simeÎncă nu există evaluări
- PinocitozaDocument26 paginiPinocitozaMonicaÎncă nu există evaluări
- LP 2 - Preparatul Microscopic PermanentDocument30 paginiLP 2 - Preparatul Microscopic PermanentMihaiLÎncă nu există evaluări
- Reglarea Exprimării Genelor La EucarioteDocument8 paginiReglarea Exprimării Genelor La EucariotePristavu Mircea-CosminÎncă nu există evaluări
- Metabolismul MicrobianDocument11 paginiMetabolismul MicrobianRazvan RastiuÎncă nu există evaluări
- Fenomene de Transport MembranarDocument23 paginiFenomene de Transport MembranarBfkt ProÎncă nu există evaluări
- Permeabilitate Şi TransportDocument14 paginiPermeabilitate Şi TransportLisa Briggs100% (1)
- Biofizica 7Document34 paginiBiofizica 7Lile George100% (1)
- Curs 4 BIOFIZDocument72 paginiCurs 4 BIOFIZfloricapopovici168Încă nu există evaluări
- WWW - Referate.ro-Membrana Celulara Structura Moleculara Si Functiile Membranelor Celulare 81e12Document25 paginiWWW - Referate.ro-Membrana Celulara Structura Moleculara Si Functiile Membranelor Celulare 81e12Otilia Iftimie100% (1)
- Membrana CelularaDocument25 paginiMembrana CelularaFlavius DobreÎncă nu există evaluări
- Colesterolul MembranarDocument5 paginiColesterolul MembranarProf. Cristina GheraÎncă nu există evaluări
- S5 Subiect PDFDocument19 paginiS5 Subiect PDFAna-Maria NănescuÎncă nu există evaluări
- Hamza Bianca - Georgiana: Curriculum VitaeDocument2 paginiHamza Bianca - Georgiana: Curriculum VitaeRaluca RoxanaÎncă nu există evaluări
- S1 SubiectDocument2 paginiS1 SubiectRaluca RoxanaÎncă nu există evaluări
- S1 SubiectDocument20 paginiS1 SubiectMihai TudorÎncă nu există evaluări
- Curs 8 Metabolismul Bilirubinei - Sindromul Icteric. Acizii Biliari 1Document41 paginiCurs 8 Metabolismul Bilirubinei - Sindromul Icteric. Acizii Biliari 1Raluca Roxana100% (1)
- Curs 10 Ca, MG, PDocument22 paginiCurs 10 Ca, MG, PGalea-Holhoș Larisa Bianca100% (1)
- 1223 Anexa 4 - 8729 - 6644Document14 pagini1223 Anexa 4 - 8729 - 6644Raluca RoxanaÎncă nu există evaluări
- Orl PDFDocument73 paginiOrl PDFRaluca RoxanaÎncă nu există evaluări
- Neuro ChirDocument75 paginiNeuro ChirRaluca RoxanaÎncă nu există evaluări
- Neuro ChirDocument75 paginiNeuro ChirRaluca RoxanaÎncă nu există evaluări
- TermeniDocument5 paginiTermeniRaluca RoxanaÎncă nu există evaluări
- WWW - Referat.ro ExpresionismulinpoeziaEunustrivescCorolademinunialumii 354f2Document9 paginiWWW - Referat.ro ExpresionismulinpoeziaEunustrivescCorolademinunialumii 354f2Raluca RoxanaÎncă nu există evaluări
- Curs Apendicita RezidentiDocument66 paginiCurs Apendicita RezidentiPavel MarianÎncă nu există evaluări
- Artere Si VeneDocument2 paginiArtere Si VeneRaluca RoxanaÎncă nu există evaluări
- Filosof Nat UmanaDocument2 paginiFilosof Nat UmanaRaluca RoxanaÎncă nu există evaluări
- Artrologie GeneralitatiDocument4 paginiArtrologie GeneralitatiCristina NuţaşÎncă nu există evaluări