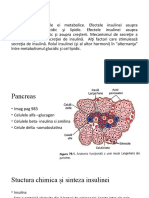
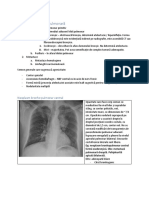
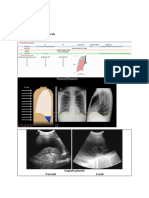
S-ar putea să vă placă și
- Metabolismul LipidicDocument35 paginiMetabolismul LipidicEmese Pop100% (4)
- C8 - Glandele SuprarenaleDocument20 paginiC8 - Glandele SuprarenaleSpoiala DragosÎncă nu există evaluări
- 3 Metabolismul Lipidic - ColesterolDocument38 pagini3 Metabolismul Lipidic - ColesterolHawah Raluca100% (3)
- Metabolismul GlucidicDocument48 paginiMetabolismul GlucidicAndreea Alida100% (3)
- Ciclul KrebsDocument28 paginiCiclul KrebstetodoraÎncă nu există evaluări
- Colesterolul - ReferatDocument7 paginiColesterolul - ReferatIoana BratuÎncă nu există evaluări
- Metabolismul Lipidelor 8 MaiDocument82 paginiMetabolismul Lipidelor 8 MaiSanda BuruianăÎncă nu există evaluări
- FiziopatologieDocument6 paginiFiziopatologieBetaÎncă nu există evaluări
- Gluconeogeneza 2022 PDFDocument11 paginiGluconeogeneza 2022 PDFLaura MunteanuÎncă nu există evaluări
- LipoproteineDocument25 paginiLipoproteineLarisa Maria IlieÎncă nu există evaluări
- Curs Vitamine HidrosolubileDocument40 paginiCurs Vitamine HidrosolubileLeonard D50% (2)
- Curs PancreasDocument17 paginiCurs PancreasZÎncă nu există evaluări
- Curs 11 Enzime Cu Semnificatie Clinica G-6-PD, GT, Lipaza, AldolazaDocument29 paginiCurs 11 Enzime Cu Semnificatie Clinica G-6-PD, GT, Lipaza, AldolazaAndrei E. HawayÎncă nu există evaluări
- Farmacoterapia DislipidemiilorDocument54 paginiFarmacoterapia DislipidemiilorLiviu LivaxÎncă nu există evaluări
- HORMONIIDocument9 paginiHORMONIIandreeamihai92Încă nu există evaluări
- Biochimie 7 - Metabolismul Glucidelor I (Glicoliza, Decarboxilarea, Ciclul Krebs) CATB (An III)Document55 paginiBiochimie 7 - Metabolismul Glucidelor I (Glicoliza, Decarboxilarea, Ciclul Krebs) CATB (An III)Alex Andru100% (2)
- Lipoproteinele PlasmaticeDocument19 paginiLipoproteinele PlasmaticeMihaela CarnutaÎncă nu există evaluări
- Metabolismul LipidicDocument21 paginiMetabolismul Lipidicmadalina31Încă nu există evaluări
- PREL V - Glucidele IDocument106 paginiPREL V - Glucidele IZmeu CristinaÎncă nu există evaluări
- Biochimie - Laura - Cap3 - Metabolism GlucidicDocument36 paginiBiochimie - Laura - Cap3 - Metabolism GlucidicMariaÎncă nu există evaluări
- HEMOPROTEINEDocument6 paginiHEMOPROTEINEGherman MadalinaÎncă nu există evaluări
- Insulina GlucagonDocument44 paginiInsulina GlucagonAlexandru IarovoiÎncă nu există evaluări
- Metabolismul Lipidelor Şi LipoproteinelorDocument9 paginiMetabolismul Lipidelor Şi LipoproteinelorcheciudianaÎncă nu există evaluări
- 5.metabolismul Glucidic - Metabolismul GlicogenuluiDocument18 pagini5.metabolismul Glucidic - Metabolismul Glicogenuluilucastefan21Încă nu există evaluări
- Metabolismul LipoproteinelorDocument49 paginiMetabolismul LipoproteinelorCiocarlan MihaiÎncă nu există evaluări
- Metabolismul LipidicDocument83 paginiMetabolismul LipidicMihaela DidencuÎncă nu există evaluări
- Biocel Sub Rezolvate 2009Document25 paginiBiocel Sub Rezolvate 2009ariadna89100% (2)
- Metabolismul Magneziului 7Document7 paginiMetabolismul Magneziului 7Diana BenchiaÎncă nu există evaluări
- 94glicoliza Şi GluconeogenezaDocument28 pagini94glicoliza Şi GluconeogenezaRalucaIoana16100% (1)
- 3 BH MD Ro Metabolism LipidicDocument51 pagini3 BH MD Ro Metabolism LipidicStef DavidÎncă nu există evaluări
- Metabolismul LipidelorDocument251 paginiMetabolismul Lipidelorgvmeareason78% (9)
- Metabolismul FructozeiDocument15 paginiMetabolismul FructozeiAnomar MRg50% (2)
- Dozarea ColesteroluluiDocument1 paginăDozarea ColesteroluluiAdina LauraÎncă nu există evaluări
- Rolul Ficatului În Biochimia LipidelorDocument6 paginiRolul Ficatului În Biochimia Lipidelorlaurisca937660Încă nu există evaluări
- Analize Medical ElaboratorDocument12 paginiAnalize Medical ElaboratorarmurereÎncă nu există evaluări
- Lucrarea de Licenta DiscutiiDocument1 paginăLucrarea de Licenta DiscutiiMihai HondruÎncă nu există evaluări
- Patologie Tumorală PulmonarăDocument5 paginiPatologie Tumorală PulmonarăMihai HondruÎncă nu există evaluări
- Metabolismul ColesteroluluiDocument42 paginiMetabolismul Colesteroluluielena petreÎncă nu există evaluări
- ProteineDocument3 paginiProteineAnnaMincaÎncă nu există evaluări
- Curs 6 Glicogen, Fru, GalDocument12 paginiCurs 6 Glicogen, Fru, GalMarkus ZahndÎncă nu există evaluări
- Curs 2 Biochimie Clinica PDFDocument9 paginiCurs 2 Biochimie Clinica PDFCristina MurgociÎncă nu există evaluări
- Curs 2 Biochimie Clinica - DZDocument9 paginiCurs 2 Biochimie Clinica - DZAlexandra Nedelcoff-GhiranlieffÎncă nu există evaluări
- GlicocalixDocument16 paginiGlicocalixivanalexandra0% (1)
- Determinarea Activitatii GGT BiochimieDocument2 paginiDeterminarea Activitatii GGT BiochimieDiana DianaDdÎncă nu există evaluări
- Metabolismul GlucidelorDocument151 paginiMetabolismul GlucidelorGura ColeaÎncă nu există evaluări
- Subiecte BiochimieDocument1 paginăSubiecte BiochimieManda AndreeaÎncă nu există evaluări
- Caragenan - E407Document9 paginiCaragenan - E407AndreeaÎncă nu există evaluări
- Curs 3 Aparatul GolgiDocument4 paginiCurs 3 Aparatul GolgiOana Koppandi100% (1)
- Calea PentozofosfatilorDocument4 paginiCalea Pentozofosfatilorwings_of_ang3lÎncă nu există evaluări
- Curs 2 Biochimie II Metab Lipidic 4Document13 paginiCurs 2 Biochimie II Metab Lipidic 4Alex AlexÎncă nu există evaluări
- Curs 5 - LipoproteineDocument24 paginiCurs 5 - LipoproteineCosmina FuturemedÎncă nu există evaluări
- Subiecte LipidicDocument11 paginiSubiecte LipidicMihai AnaÎncă nu există evaluări
- Lipoproteine PlasmaticeDocument13 paginiLipoproteine PlasmaticeipaperrrÎncă nu există evaluări
- Lipoproteinele PlasmaticeDocument9 paginiLipoproteinele PlasmaticeAna MariaÎncă nu există evaluări
- FP Metab. LipidicDocument13 paginiFP Metab. LipidicBuggieÎncă nu există evaluări
- Lipoproteinele PlasmaticeDocument22 paginiLipoproteinele PlasmaticeCatalin MihaiÎncă nu există evaluări
- Metabolism Lipidic PDFDocument51 paginiMetabolism Lipidic PDFDudu AliceÎncă nu există evaluări
- ZZZZZDocument185 paginiZZZZZalla7maraqaÎncă nu există evaluări
- Metabolism Lipidic MDDocument110 paginiMetabolism Lipidic MDElena CristinaÎncă nu există evaluări
- CIRCUITUL METABOLIC AL LIPOPROTEINELOR StudentiDocument7 paginiCIRCUITUL METABOLIC AL LIPOPROTEINELOR StudentiAnanya PaunÎncă nu există evaluări
- Metabolismul LipidicDocument37 paginiMetabolismul LipidicArina ChifulescuÎncă nu există evaluări
- HiperlipoproteinemiiDocument4 paginiHiperlipoproteinemiiRujan Marcel-adrianÎncă nu există evaluări
- Bronșiectazii, BPOC, Sarcoidoză Și PneumoconiozeDocument7 paginiBronșiectazii, BPOC, Sarcoidoză Și PneumoconiozeMihai HondruÎncă nu există evaluări
- Patologie PleuralăDocument10 paginiPatologie PleuralăMihai HondruÎncă nu există evaluări
- MediastinulDocument9 paginiMediastinulMihai HondruÎncă nu există evaluări
- Cardio VascularDocument15 paginiCardio VascularMihai HondruÎncă nu există evaluări
- 1SUBIECTE - Tematica Examen Oncologie ROM 2020Document4 pagini1SUBIECTE - Tematica Examen Oncologie ROM 2020Mihai HondruÎncă nu există evaluări
- IntroducereDocument10 paginiIntroducereMihai HondruÎncă nu există evaluări
- Explorarea Câmpului Vizual - LP StudențiDocument25 paginiExplorarea Câmpului Vizual - LP StudențiMihai HondruÎncă nu există evaluări
- Prezentare PIO StudentiDocument26 paginiPrezentare PIO StudentiMihai HondruÎncă nu există evaluări
- Simțul Cromatic Examen NicolDocument37 paginiSimțul Cromatic Examen NicolMihai HondruÎncă nu există evaluări
- Determinarea PioDocument1 paginăDeterminarea PioMihai HondruÎncă nu există evaluări
- Diagnosticul Diferențial Al Ochiului Roșu NicolDocument22 paginiDiagnosticul Diferențial Al Ochiului Roșu NicolMihai HondruÎncă nu există evaluări
- Simțul CromaticDocument37 paginiSimțul CromaticMihai HondruÎncă nu există evaluări
- Hernii Anterolaterale AbdomenDocument2 paginiHernii Anterolaterale AbdomenMihai HondruÎncă nu există evaluări
- Proiect CalcitoninaDocument4 paginiProiect CalcitoninaMihai HondruÎncă nu există evaluări
- AnemieDocument2 paginiAnemieMihai HondruÎncă nu există evaluări
- MorfopatologieDocument6 paginiMorfopatologieMihai HondruÎncă nu există evaluări
- SemioChir 2Document2 paginiSemioChir 2Mihai HondruÎncă nu există evaluări
- Lucrarea de Licenta Reguli Pentru Redactarea Tabelelor Si FigurilorDocument1 paginăLucrarea de Licenta Reguli Pentru Redactarea Tabelelor Si FigurilorMihai HondruÎncă nu există evaluări
- TraheostomieDocument13 paginiTraheostomieMihai HondruÎncă nu există evaluări