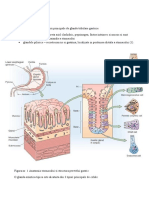
S-ar putea să vă placă și
- Gluconeogeneza 2022 PDFDocument11 paginiGluconeogeneza 2022 PDFLaura MunteanuÎncă nu există evaluări
- 5 - Digestia Glucidelor - Metabolismul GlicogenluiDocument18 pagini5 - Digestia Glucidelor - Metabolismul Glicogenluinagyantonia100% (1)
- Biochimie SintezeDocument27 paginiBiochimie SintezeChristian DanielÎncă nu există evaluări
- HEMOPROTEINEDocument6 paginiHEMOPROTEINEGherman MadalinaÎncă nu există evaluări
- Metabolismul ProteinelorDocument12 paginiMetabolismul ProteinelorCalin-Sorin CimpeanuÎncă nu există evaluări
- Metablismul NucleotidelorDocument60 paginiMetablismul NucleotidelorAlice DouglasÎncă nu există evaluări
- Metabolismul GlucidicDocument2 paginiMetabolismul GlucidicNica AndraÎncă nu există evaluări
- Metabolismul Protidic: Determinarea Proteinelor Totale Din Lichidele Biologice La EcvineDocument13 paginiMetabolismul Protidic: Determinarea Proteinelor Totale Din Lichidele Biologice La EcvineAlina VulpeÎncă nu există evaluări
- 14.hormoni Mecanisme Actiune, Catecolamine, Glucagon CFILIPDocument15 pagini14.hormoni Mecanisme Actiune, Catecolamine, Glucagon CFILIPLarisa ElenaÎncă nu există evaluări
- Calreticulin BioDocument18 paginiCalreticulin BioChitic VirginiaÎncă nu există evaluări
- Transcrierea AdnDocument21 paginiTranscrierea AdnPristavu Mircea-CosminÎncă nu există evaluări
- 9 ColesterolDocument138 pagini9 ColesterolMSÎncă nu există evaluări
- Curs 8 Codul GeneticDocument11 paginiCurs 8 Codul GeneticThisisme258Încă nu există evaluări
- Colesterolul MembranarDocument5 paginiColesterolul MembranarProf. Cristina GheraÎncă nu există evaluări
- Curs BiochimieDocument6 paginiCurs BiochimieMariana SandorÎncă nu există evaluări
- Biosinteza AminoacizilorDocument2 paginiBiosinteza AminoacizilorIonut NealcosÎncă nu există evaluări
- NUCLEOTIDE - STDocument27 paginiNUCLEOTIDE - STCristiana AndreeaÎncă nu există evaluări
- Proteine Plasmatice Curs PuDocument9 paginiProteine Plasmatice Curs PuGabrielaÎncă nu există evaluări
- Calea Pentozofosfat - Calea UronicaDocument12 paginiCalea Pentozofosfat - Calea UronicaAlex DanciuÎncă nu există evaluări
- Hemoglobina GlicozilatăDocument2 paginiHemoglobina GlicozilatăJohnny Corday100% (1)
- Curs 3 GNG-C. Pentozo-Fosfat - C. Glucuronica 2017-FinalDocument62 paginiCurs 3 GNG-C. Pentozo-Fosfat - C. Glucuronica 2017-FinalPopa StefanaÎncă nu există evaluări
- NucleotideDocument30 paginiNucleotideCrystyan Cryss100% (1)
- Rinichiul ArtificialDocument5 paginiRinichiul Artificiallucscurtu100% (1)
- Curs 1-Imunogenetica Si Transplantul - ComplexulDocument33 paginiCurs 1-Imunogenetica Si Transplantul - ComplexulCristian EneÎncă nu există evaluări
- Lantul RespiratorDocument80 paginiLantul RespiratorElena MunteanuÎncă nu există evaluări
- Homeostazia Apei Si A SodiuluiDocument5 paginiHomeostazia Apei Si A SodiuluiAlyna Marya IoaneiÎncă nu există evaluări
- CURS 11 Metabolismul Lipidelor 1Document60 paginiCURS 11 Metabolismul Lipidelor 1waseemÎncă nu există evaluări
- Sistemul Nervos Vegetativ - COLINERGICDocument14 paginiSistemul Nervos Vegetativ - COLINERGICMihai MarinÎncă nu există evaluări
- 1b Lanțul Transportor de ElectroniDocument82 pagini1b Lanțul Transportor de ElectroniLaura HutzelÎncă nu există evaluări
- Metabolism LipideDocument112 paginiMetabolism LipideAlexandra Ioana MoatarÎncă nu există evaluări
- SulfanilamideDocument15 paginiSulfanilamidejalbucristinaÎncă nu există evaluări
- Curs 2 - EritrocitulDocument27 paginiCurs 2 - EritrocitulAndreeaAllexandraÎncă nu există evaluări
- Hormonii Implicati in Mentinerea Echilibrului Fosfo Cu DiacriticeDocument9 paginiHormonii Implicati in Mentinerea Echilibrului Fosfo Cu Diacriticeniko55555Încă nu există evaluări
- Ii. Curs 4Document20 paginiIi. Curs 4Tiggy TigÎncă nu există evaluări
- Curs 11 Enzime Cu Semnificatie Clinica G-6-PD, GT, Lipaza, AldolazaDocument29 paginiCurs 11 Enzime Cu Semnificatie Clinica G-6-PD, GT, Lipaza, AldolazaAndrei E. HawayÎncă nu există evaluări
- Calea UronicaDocument7 paginiCalea UronicaSebi RaulÎncă nu există evaluări
- Metabolismul GliceroluluiDocument23 paginiMetabolismul GliceroluluiCristina Diaconu100% (1)
- Insulina GlucagonDocument44 paginiInsulina GlucagonAlexandru IarovoiÎncă nu există evaluări
- Fiziologie LP NR 6 Anul IDocument17 paginiFiziologie LP NR 6 Anul IAnna MariaÎncă nu există evaluări
- Metabolismul LipidelorDocument40 paginiMetabolismul LipidelorEma Virga100% (1)
- Enzime Si IzoenzimeDocument10 paginiEnzime Si IzoenzimeRaluca AnghelÎncă nu există evaluări
- FiziologieDocument33 paginiFiziologieRoman Mihai100% (1)
- Ureea Reprezinta Produsul Final Al Catabolismului Azotului ProteicDocument2 paginiUreea Reprezinta Produsul Final Al Catabolismului Azotului ProteicOana PaladeÎncă nu există evaluări
- C1-Biochimie Metabolică Și DescriptivăDocument6 paginiC1-Biochimie Metabolică Și DescriptivăGheorghe ElenaÎncă nu există evaluări
- Metabolism Lipidic MDDocument110 paginiMetabolism Lipidic MDElena CristinaÎncă nu există evaluări
- Calciu Mesager Secund - Put IoanaDocument17 paginiCalciu Mesager Secund - Put IoanaIoana PutÎncă nu există evaluări
- Capitolul V. Formarea Methemoglobinei Sub Acţiunea Ionilor Nitriţi PDFDocument20 paginiCapitolul V. Formarea Methemoglobinei Sub Acţiunea Ionilor Nitriţi PDFAaron WilsonÎncă nu există evaluări
- FIZIOLOGIE Sangele2Document63 paginiFIZIOLOGIE Sangele2Mihaela VadanaÎncă nu există evaluări
- Curs 6 - MitocondriaDocument17 paginiCurs 6 - MitocondriaRebecca Sandu100% (1)
- Sinteza AGDocument71 paginiSinteza AGConstantin UntilaÎncă nu există evaluări
- Metabolismul Acidului Arahidonic, EicosanoiziiDocument29 paginiMetabolismul Acidului Arahidonic, EicosanoiziiIrina AndreeaÎncă nu există evaluări
- Curs 11 Biochimie - Metab. Glucidic 4Document11 paginiCurs 11 Biochimie - Metab. Glucidic 4Claudia CojocariuÎncă nu există evaluări
- Sunt GlicogenDocument55 paginiSunt GlicogenDiana Didi DiaÎncă nu există evaluări
- Tema 9 Metabolismul Glicogenului-87706Document50 paginiTema 9 Metabolismul Glicogenului-87706Andreea SpataruÎncă nu există evaluări
- Glucide MetabolismDocument14 paginiGlucide MetabolismFlorentina PanfilÎncă nu există evaluări
- Metabolismul GlucidelorDocument7 paginiMetabolismul GlucidelorCatrinel Grăjdeanu-PetroviciÎncă nu există evaluări
- Curs 10 Biochimie - Metab. Glucidic 3Document13 paginiCurs 10 Biochimie - Metab. Glucidic 3TGÎncă nu există evaluări
- Anabolismul Glucidelor MgiDocument11 paginiAnabolismul Glucidelor Mgimari SÎncă nu există evaluări
- C6Document27 paginiC6Malina IoanaÎncă nu există evaluări
- Gli Co Geno LizaDocument3 paginiGli Co Geno LizaMiruna IoanÎncă nu există evaluări
- Plan de Afaceri Cu PEST Si SWOTDocument23 paginiPlan de Afaceri Cu PEST Si SWOTBeatrice ElenaÎncă nu există evaluări
- Management - Risc Curs 6 ISO 31000 - 2021Document40 paginiManagement - Risc Curs 6 ISO 31000 - 2021lucastefan21Încă nu există evaluări
- Managementul Riscurilor Curs 7 Riscul Subst Peric 2021Document48 paginiManagementul Riscurilor Curs 7 Riscul Subst Peric 2021lucastefan21Încă nu există evaluări
- PracticaDocument10 paginiPracticalucastefan21Încă nu există evaluări
- Proiect - Etica Si Integritate AcademicaDocument29 paginiProiect - Etica Si Integritate Academicalucastefan21100% (2)
- Manag Riscuri Curs 3 Identif Riscurilor 2021Document33 paginiManag Riscuri Curs 3 Identif Riscurilor 2021lucastefan21Încă nu există evaluări
- Manag-Risc Curs 5 Riscuri Agricult Ind Aliment 2021Document35 paginiManag-Risc Curs 5 Riscuri Agricult Ind Aliment 2021lucastefan21Încă nu există evaluări
- Istoria AudituluiDocument4 paginiIstoria Audituluilucastefan21Încă nu există evaluări
- Metabolismul Proteinelor, Nucleotidelor, CromoproteinelorDocument258 paginiMetabolismul Proteinelor, Nucleotidelor, Cromoproteinelorlucastefan21Încă nu există evaluări
- Metabolismul LipidicDocument148 paginiMetabolismul LipidicSimonaGrigorașÎncă nu există evaluări
- VitamineleDocument37 paginiVitamineleCarmenMariutÎncă nu există evaluări
- Metabolismul Lipidic - Met ColesteroluluiDocument49 paginiMetabolismul Lipidic - Met Colesteroluluilucastefan21Încă nu există evaluări
- MANAGEMENTUL RISCURILOR Curs 11 Manag Riscuri AprovizionareDocument39 paginiMANAGEMENTUL RISCURILOR Curs 11 Manag Riscuri Aprovizionarelucastefan21Încă nu există evaluări
- Auditarea Situatiilor FinanciareDocument11 paginiAuditarea Situatiilor Financiarelucastefan21Încă nu există evaluări
- Fiziologie - CursDocument29 paginiFiziologie - CursGeorgiana SpatariuÎncă nu există evaluări
- VitamineleDocument37 paginiVitamineleCarmenMariutÎncă nu există evaluări
- ProteineleDocument20 paginiProteineleVinitinschi MaximÎncă nu există evaluări
- PolarimetrieDocument7 paginiPolarimetrielucastefan21100% (1)
- Boala ScafandruluiDocument7 paginiBoala Scafandruluilucastefan21Încă nu există evaluări
- 5.metabolismul Glucidic - Metabolismul GlicogenuluiDocument18 pagini5.metabolismul Glucidic - Metabolismul Glicogenuluilucastefan21Încă nu există evaluări
- Proiect - Psihologie MedicalaDocument4 paginiProiect - Psihologie Medicalalucastefan21Încă nu există evaluări
- Pct. 2 Situatii Financiare La 31.12.2019RODocument3 paginiPct. 2 Situatii Financiare La 31.12.2019ROlucastefan21Încă nu există evaluări
- Contract de Munca - Model ITMDocument2 paginiContract de Munca - Model ITMdanileonteÎncă nu există evaluări
- Id102 - Raportul Auditorului Financiar 2004Document12 paginiId102 - Raportul Auditorului Financiar 2004lucastefan21Încă nu există evaluări
- Aprob: R O M Â N I A Ministerul Afacerilor Interne Anexa Nr. 10 Nesecret Bucureşti Nr. 150.393 Din 30.09.2016 Ex. UnicDocument4 paginiAprob: R O M Â N I A Ministerul Afacerilor Interne Anexa Nr. 10 Nesecret Bucureşti Nr. 150.393 Din 30.09.2016 Ex. UnicAdy PastorÎncă nu există evaluări
- Curs Id Co-2019 IV IeaDocument99 paginiCurs Id Co-2019 IV Iealucastefan21Încă nu există evaluări
- Ghid Finalizare StudiiDocument11 paginiGhid Finalizare StudiiYvonne IoanaÎncă nu există evaluări
- Marketing International (Constantin Sasu)Document185 paginiMarketing International (Constantin Sasu)Marina Gindac100% (1)
- 1 PDFDocument4 pagini1 PDFIacob Cristian-AlexandruÎncă nu există evaluări