S-ar putea să vă placă și
- PREL V - Glucidele IDocument106 paginiPREL V - Glucidele IZmeu CristinaÎncă nu există evaluări
- Gluconeogeneza 2022 PDFDocument11 paginiGluconeogeneza 2022 PDFLaura MunteanuÎncă nu există evaluări
- Curs 4 Metabolismul GlucidelorDocument11 paginiCurs 4 Metabolismul Glucideloraliaj2Încă nu există evaluări
- Gli Co Geno LizaDocument3 paginiGli Co Geno LizaMiruna IoanÎncă nu există evaluări
- Metab. Glucidic - Curs 1, BiochimieDocument46 paginiMetab. Glucidic - Curs 1, BiochimieGreta IulianaÎncă nu există evaluări
- Functia ReflexaDocument3 paginiFunctia ReflexaAna AlexăndroaeÎncă nu există evaluări
- ProteineDocument3 paginiProteineAnnaMincaÎncă nu există evaluări
- DIABETDocument34 paginiDIABETMarcu SanzianaÎncă nu există evaluări
- C1-Biochimie Metabolică Și DescriptivăDocument6 paginiC1-Biochimie Metabolică Și DescriptivăGheorghe ElenaÎncă nu există evaluări
- EnzimologieDocument6 paginiEnzimologieRoxana BoncoiÎncă nu există evaluări
- Glicoliza PiruvicDocument15 paginiGlicoliza PiruvicTărnăuceanu Cosmin ConstantinÎncă nu există evaluări
- Lipidele & AntioxidantiiDocument20 paginiLipidele & AntioxidantiiDaniel Barta100% (1)
- Nutritie Anul 2Document5 paginiNutritie Anul 2Moruzi Anamaria100% (1)
- Fermentatia Propionica ProiectDocument3 paginiFermentatia Propionica ProiectMorosan DanaÎncă nu există evaluări
- C1 Metabolism Generalitati 2021Document39 paginiC1 Metabolism Generalitati 2021gabriela serban100% (1)
- Genetica CursDocument4 paginiGenetica CursRonaldo LazarÎncă nu există evaluări
- Catabolismul TrigliceridelorDocument21 paginiCatabolismul Trigliceridelorcristinacristina13Încă nu există evaluări
- Metabolismul Magneziului 7Document7 paginiMetabolismul Magneziului 7Diana BenchiaÎncă nu există evaluări
- LipazeleDocument11 paginiLipazelealxo91Încă nu există evaluări
- Metabolism Corpi Cetonici & Lipide ComplexeDocument10 paginiMetabolism Corpi Cetonici & Lipide ComplexeSilvana AlbertÎncă nu există evaluări
- Metabolismul Lipidelor 8 MaiDocument82 paginiMetabolismul Lipidelor 8 MaiSanda BuruianăÎncă nu există evaluări
- Nutritia MicroorganismelorDocument6 paginiNutritia MicroorganismelorDana GeorgianaÎncă nu există evaluări
- Fiziologia Muschilor - Point PresentationDocument58 paginiFiziologia Muschilor - Point PresentationmariusÎncă nu există evaluări
- EnzimeleDocument3 paginiEnzimeleGeorgiana ZahariaÎncă nu există evaluări
- Sistemul NervosDocument20 paginiSistemul NervosPaviliuc Raluca100% (1)
- Subiecte BiochimieDocument1 paginăSubiecte BiochimieManda AndreeaÎncă nu există evaluări
- CELULADocument18 paginiCELULASabina PostolacheÎncă nu există evaluări
- CalciuDocument67 paginiCalciulilikauzun88100% (1)
- Receptori BiochimieDocument96 paginiReceptori Biochimiedeynyk0% (1)
- SUBIECTE Fiziologie An 2 Sem 2 Pe ScurtDocument71 paginiSUBIECTE Fiziologie An 2 Sem 2 Pe ScurtPatricia Ariana100% (1)
- GlumatDocument13 paginiGlumatGabriela TudoranÎncă nu există evaluări
- Catabolismul Hemului. Bilirubina.Document5 paginiCatabolismul Hemului. Bilirubina.Vasilache AlinaÎncă nu există evaluări
- Curs 2 Clasificare. HemoglobinaDocument20 paginiCurs 2 Clasificare. Hemoglobinajacksparrow2Încă nu există evaluări
- C14.1 GlucocorticoiziiDocument20 paginiC14.1 GlucocorticoiziiGeo GrigoreÎncă nu există evaluări
- Sinteza Si Secretia CelularaDocument6 paginiSinteza Si Secretia Celularaabcfree2006Încă nu există evaluări
- Curs Metab. Glucidic - 2Document10 paginiCurs Metab. Glucidic - 2Onofrei MariaÎncă nu există evaluări
- Lactat Dehidrogeneza (LDH) - Izoenzime - SynevoDocument4 paginiLactat Dehidrogeneza (LDH) - Izoenzime - SynevoLaurentiu Dinu100% (1)
- C8 Metabolism PROTEINE IPA 20161Document37 paginiC8 Metabolism PROTEINE IPA 20161Bogdan NaeÎncă nu există evaluări
- Testul PCRDocument9 paginiTestul PCRIulia BrindescuÎncă nu există evaluări
- 0 FotosintezaDocument8 pagini0 FotosintezaDorin BorshÎncă nu există evaluări
- Curs 3Document30 paginiCurs 3Anamaria PetrutÎncă nu există evaluări
- Caragenan - E407Document9 paginiCaragenan - E407AndreeaÎncă nu există evaluări
- 3 BH MD Ro Metabolism LipidicDocument51 pagini3 BH MD Ro Metabolism LipidicStef DavidÎncă nu există evaluări
- GlucideleDocument25 paginiGlucideleMasha GorasÎncă nu există evaluări
- StereoizomerieDocument10 paginiStereoizomerieFelicia VerdesÎncă nu există evaluări
- Curs 2 Biochimie Clinica PDFDocument9 paginiCurs 2 Biochimie Clinica PDFCristina MurgociÎncă nu există evaluări
- UreazaDocument10 paginiUreazaAlexandra BurlacuÎncă nu există evaluări
- Curs 2 Biochimie II - Metab. Lipidic 4Document12 paginiCurs 2 Biochimie II - Metab. Lipidic 4Mihai HondruÎncă nu există evaluări
- Lizozomi Peroxizomi 2018Document106 paginiLizozomi Peroxizomi 2018Iulia Talpalariu100% (1)
- Metabolismul LipoproteinelorDocument49 paginiMetabolismul LipoproteinelorCiocarlan MihaiÎncă nu există evaluări
- Dismetabolisme LipiiceDocument20 paginiDismetabolisme LipiicePavel GaladiucÎncă nu există evaluări
- FIZIOLOGIE Sangele2Document63 paginiFIZIOLOGIE Sangele2Mihaela VadanaÎncă nu există evaluări
- Curs 5 BiochimieDocument52 paginiCurs 5 Biochimiemihaela lovinÎncă nu există evaluări
- Tema 9 Metabolismul Glicogenului-87706Document50 paginiTema 9 Metabolismul Glicogenului-87706Andreea SpataruÎncă nu există evaluări
- Metabolismul Glucidic An I AMGDocument15 paginiMetabolismul Glucidic An I AMGMiulescu CristinaÎncă nu există evaluări
- Catabolism Si AnabolismDocument20 paginiCatabolism Si AnabolismBogdan CaliniucÎncă nu există evaluări
- Curs 11 Biochimie - Metab. Glucidic 4Document11 paginiCurs 11 Biochimie - Metab. Glucidic 4Claudia CojocariuÎncă nu există evaluări
- GlucideDocument5 paginiGlucideeugeniu222Încă nu există evaluări
- Anabolismul Glucidelor MgiDocument11 paginiAnabolismul Glucidelor Mgimari SÎncă nu există evaluări
- Prezentare Metabolismul GlucidelorDocument18 paginiPrezentare Metabolismul GlucidelorMarina Denisa Ilina100% (1)
- Human AnatomyDocument14 paginiHuman AnatomyMarkus ZahndÎncă nu există evaluări
- Portofoliu Activitati Practice Biochimie-1 LPDocument14 paginiPortofoliu Activitati Practice Biochimie-1 LPMarkus ZahndÎncă nu există evaluări
- 3.morfologie Arcade DentareDocument6 pagini3.morfologie Arcade DentareMarkus ZahndÎncă nu există evaluări
- Artera FemuralăDocument3 paginiArtera FemuralăMarkus ZahndÎncă nu există evaluări
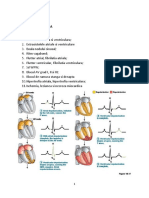
- Lucrari Practice 2 Sem 2 Electrocardiografie 2Document51 paginiLucrari Practice 2 Sem 2 Electrocardiografie 2Markus ZahndÎncă nu există evaluări
- Endo BazaDocument5 paginiEndo BazaMarkus ZahndÎncă nu există evaluări
- Fiziologie Anul I Electrocardiografie 2 + 3Document49 paginiFiziologie Anul I Electrocardiografie 2 + 3Markus ZahndÎncă nu există evaluări
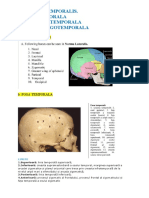
- Nortma Temporalis. Fosa Temporala Fosa Infratemporala Fosa PterigotemporalaDocument4 paginiNortma Temporalis. Fosa Temporala Fosa Infratemporala Fosa PterigotemporalaMarkus ZahndÎncă nu există evaluări
- Nervul Olftalmic. Nervul Maxilar. Nervul Mandibular Ganglionii Ciliar, Pterigopalatin, Otic, Submandibular Glandele Submandibulara Si SublingualaDocument3 paginiNervul Olftalmic. Nervul Maxilar. Nervul Mandibular Ganglionii Ciliar, Pterigopalatin, Otic, Submandibular Glandele Submandibulara Si SublingualaMarkus ZahndÎncă nu există evaluări
- Totanato LPDocument30 paginiTotanato LPMarkus ZahndÎncă nu există evaluări
- Orbita - Cav NazalaDocument5 paginiOrbita - Cav NazalaMarkus ZahndÎncă nu există evaluări
- Curs FiziologieDocument34 paginiCurs FiziologieMarkus ZahndÎncă nu există evaluări
- Curs 13 Biochimia DintilorDocument26 paginiCurs 13 Biochimia DintilorMarkus ZahndÎncă nu există evaluări
- Biochimie LP - IMCDocument7 paginiBiochimie LP - IMCMarcosÎncă nu există evaluări
- MUSCHIIDocument6 paginiMUSCHIIMarkus ZahndÎncă nu există evaluări
- Muschii Masticatori ExamenDocument4 paginiMuschii Masticatori ExamenMarkus ZahndÎncă nu există evaluări
- Anatomie LP Regiunea AxilaraDocument6 paginiAnatomie LP Regiunea AxilaraMarkus ZahndÎncă nu există evaluări
- Exo BazaDocument2 paginiExo BazaMarkus ZahndÎncă nu există evaluări
- Sfaturi BobociDocument2 paginiSfaturi BobociMarkus ZahndÎncă nu există evaluări
- Endo BazaDocument5 paginiEndo BazaMarkus ZahndÎncă nu există evaluări
- Curs 12 Hemul Degradare Si BiosintezaDocument16 paginiCurs 12 Hemul Degradare Si BiosintezaMarkus ZahndÎncă nu există evaluări
- Curs 7 Metabolism LipideDocument15 paginiCurs 7 Metabolism LipideMarkus ZahndÎncă nu există evaluări
- Curs 8 Biosinteza AG, ColesterolDocument13 paginiCurs 8 Biosinteza AG, ColesterolMarkus ZahndÎncă nu există evaluări
- Curs 5 PetozoP, Ac GlucuronicDocument10 paginiCurs 5 PetozoP, Ac GlucuronicMarkus ZahndÎncă nu există evaluări