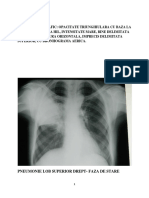
S-ar putea să vă placă și
- Cordul Pulmonar CronicDocument14 paginiCordul Pulmonar CronicAlexandra BabeanuÎncă nu există evaluări
- Explorarea Functiei RespiratoriiDocument4 paginiExplorarea Functiei RespiratoriiMoldoveanu NicoletaÎncă nu există evaluări
- Prof. Univ - DR - Egidia Miftode Tranzitia de La Terapia Antimicrobiana Intravenoasa La Cea OralaDocument21 paginiProf. Univ - DR - Egidia Miftode Tranzitia de La Terapia Antimicrobiana Intravenoasa La Cea OralapaulÎncă nu există evaluări
- Prezentare Caz BPOC 1Document6 paginiPrezentare Caz BPOC 1CrinaÎncă nu există evaluări
- 6 .Curs Bronsiolita 2014 PDFDocument65 pagini6 .Curs Bronsiolita 2014 PDFRoxanaÎncă nu există evaluări
- 6 PneumoniiDocument43 pagini6 Pneumoniicarmen_bute7337Încă nu există evaluări
- Intubarea SelectivaDocument2 paginiIntubarea Selectivasergu_20Încă nu există evaluări
- Testul de Efort EcgDocument7 paginiTestul de Efort EcgIonescu carmenÎncă nu există evaluări
- Fiziologia Respiratiei - RezumatDocument4 paginiFiziologia Respiratiei - RezumatVlad TaralaÎncă nu există evaluări
- Bacili Gram NegativDocument20 paginiBacili Gram Negativili_eÎncă nu există evaluări
- QRS Supraventricular Si QRS Ventricular (Algoritm Brugada)Document1 paginăQRS Supraventricular Si QRS Ventricular (Algoritm Brugada)Cecilia Durdui100% (1)
- 1 Fiziopatologie Astm BronsicDocument1 pagină1 Fiziopatologie Astm BronsicIrina AdrianaÎncă nu există evaluări
- Tuberculoza PulmonaraDocument8 paginiTuberculoza PulmonaraAlex SÎncă nu există evaluări
- Criterii de Clasificare Si Activitate ARDocument40 paginiCriterii de Clasificare Si Activitate AROana Andreea UrsicăÎncă nu există evaluări
- RX Studenti PlamanDocument39 paginiRX Studenti PlamanIrina AdrianaÎncă nu există evaluări
- Măsurarea Volumelor Pulmonare StaticeDocument4 paginiMăsurarea Volumelor Pulmonare StaticeRareş SecarăÎncă nu există evaluări
- Traheobronsita Acuta Si BronsiectaziileDocument29 paginiTraheobronsita Acuta Si BronsiectaziileMARANNAAÎncă nu există evaluări
- Resp 2 2019-2020Document41 paginiResp 2 2019-2020Xerox medÎncă nu există evaluări
- Curs BPOC 2019Document98 paginiCurs BPOC 2019Bianca PercsiÎncă nu există evaluări
- Sarcoidoza (Harrison)Document7 paginiSarcoidoza (Harrison)Camelia CameÎncă nu există evaluări
- Limfadenita MezentericăDocument5 paginiLimfadenita MezentericăDaniella DenissaÎncă nu există evaluări
- Fiziopatologia Insuficientei de OxigenDocument64 paginiFiziopatologia Insuficientei de OxigenLaura DianaÎncă nu există evaluări
- AnemiiDocument21 paginiAnemiiȘtefan SpînuÎncă nu există evaluări
- Curs 7 Bronhopneumopatia Obstructiva CronicaDocument39 paginiCurs 7 Bronhopneumopatia Obstructiva CronicaIosif Crin BorșoșÎncă nu există evaluări
- Tuberculoza Asociata SarciniiDocument13 paginiTuberculoza Asociata SarciniiClaudia IulianaÎncă nu există evaluări
- 6 Osteocondrita Capului FemuralDocument15 pagini6 Osteocondrita Capului FemuralMihaela IonÎncă nu există evaluări
- Capitolul 4 - RespiratorDocument34 paginiCapitolul 4 - RespiratorDiana-Elena SavaÎncă nu există evaluări
- 13484-Pielonefrita Acută La CopilDocument2 pagini13484-Pielonefrita Acută La CopilLiliana Mitu PanfiliiÎncă nu există evaluări
- Sindromul HellpDocument10 paginiSindromul HellpOana BalintÎncă nu există evaluări
- Pneumotoraxul Spontan La Nou NascutDocument2 paginiPneumotoraxul Spontan La Nou NascutAnonymous nckmiqÎncă nu există evaluări
- Curs Investigatii in PediatrieDocument51 paginiCurs Investigatii in PediatrieTiberiu-o CodrinÎncă nu există evaluări
- IntubatiaDocument4 paginiIntubatiakanzia19845166Încă nu există evaluări
- Aparatul RespiratorDocument17 paginiAparatul RespiratoryepoorikÎncă nu există evaluări
- Pulmonologie 4-5-6Document107 paginiPulmonologie 4-5-6Liuba AnghelÎncă nu există evaluări
- GSFS Studenti 2013Document48 paginiGSFS Studenti 2013ada9002Încă nu există evaluări
- Clasificarea TumorilorDocument13 paginiClasificarea Tumorilorbuzzy12Încă nu există evaluări
- Maduva SpinariiDocument2 paginiMaduva SpinariiRadu CataniciuÎncă nu există evaluări
- Managementul Cailor Aeriene Si VentilatiaDocument82 paginiManagementul Cailor Aeriene Si VentilatiaClaudia Bianca OlteanÎncă nu există evaluări
- Ciornei Monica - Fiziopatologie BPOCDocument14 paginiCiornei Monica - Fiziopatologie BPOCMonica CiorneiÎncă nu există evaluări
- Fibroscopia BronsicaDocument2 paginiFibroscopia BronsicabaczonifÎncă nu există evaluări
- Apnee CursDocument30 paginiApnee CursMihaela Gabriela RaduÎncă nu există evaluări
- Astmul Bronşic La Sugar Şi Copilul MicDocument17 paginiAstmul Bronşic La Sugar Şi Copilul MicadrianaÎncă nu există evaluări
- Sindromul Hepato-PulmonarDocument11 paginiSindromul Hepato-PulmonarMaxim ŢarălungăÎncă nu există evaluări
- 0 Reacţii de Hipersensibilitate La AINSDocument4 pagini0 Reacţii de Hipersensibilitate La AINSdregleavÎncă nu există evaluări
- Anatomia Ap RespiratorDocument10 paginiAnatomia Ap RespiratorAndreea AfrăsineiÎncă nu există evaluări
- Echinococcoza PulmonarăDocument31 paginiEchinococcoza Pulmonarăsabje20102010100% (1)
- Anemia MegaloblasticaDocument29 paginiAnemia MegaloblasticaLuciana MioaraÎncă nu există evaluări
- Curs IV ASTMUL BRONSIC PDFDocument11 paginiCurs IV ASTMUL BRONSIC PDFIris Druț-PerianÎncă nu există evaluări
- Help SindromDocument29 paginiHelp SindromStegarescu PavelÎncă nu există evaluări
- Cap 12 Chir Gen - de ListatDocument264 paginiCap 12 Chir Gen - de ListatRaduÎncă nu există evaluări
- BronsiolitaDocument9 paginiBronsiolitamihalachemirelaÎncă nu există evaluări
- Sindrom ParenchimatosDocument61 paginiSindrom ParenchimatosElena123Încă nu există evaluări
- Insuficienta RespiratorieDocument41 paginiInsuficienta RespiratorieAhmad Al ShdefatÎncă nu există evaluări
- Imunitatea InnascutaDocument5 paginiImunitatea InnascutaMaria CloșcăÎncă nu există evaluări
- Curs 06. SI Nespecific - Mecanismele PasiveDocument3 paginiCurs 06. SI Nespecific - Mecanismele PasiveAndrei StefanÎncă nu există evaluări
- Introducere in ImunologieDocument20 paginiIntroducere in ImunologieAndrea Dea100% (1)
- ImunobiologieDocument44 paginiImunobiologieKfa DkhÎncă nu există evaluări
- ImunologieDocument263 paginiImunologieIlinca RizeaÎncă nu există evaluări
- Imunologie LPDocument9 paginiImunologie LPLUÎncă nu există evaluări
- ImunitateaDocument6 paginiImunitateaLuminita Brinza100% (1)
- Lavaje EndodonticeDocument18 paginiLavaje EndodonticeGeorgiana Alexandra CerneaÎncă nu există evaluări
- Ghid Redactare Lucrare LicentaDocument4 paginiGhid Redactare Lucrare LicentaVladimir DrăgoiÎncă nu există evaluări
- Lavaje EndodonticeDocument18 paginiLavaje EndodonticeGeorgiana Alexandra CerneaÎncă nu există evaluări
- Curs ManagementDocument81 paginiCurs ManagementSara HmÎncă nu există evaluări
- Curs Lucrul in EchipaDocument26 paginiCurs Lucrul in EchipaAlexandra AlexandraÎncă nu există evaluări
- Compendiu de Chirurgie OMF, Prof. Bucur, Vol IIDocument519 paginiCompendiu de Chirurgie OMF, Prof. Bucur, Vol IISergiu Si Renata Bancos100% (55)
- FizioterapieDocument223 paginiFizioterapieIoana Harpa86% (7)
- Radiologie CsaDocument4 paginiRadiologie Csakty_Încă nu există evaluări
- Insuficienta HepaticaDocument77 paginiInsuficienta HepaticaAndreea MireaÎncă nu există evaluări
- FizioterapieDocument223 paginiFizioterapieIoana Harpa86% (7)
- Carte Medicina Interna - Varianta CompletaDocument261 paginiCarte Medicina Interna - Varianta CompletaAndra Lazar71% (7)
- Radiologie Pentru Studenti Si Medici StomatologiDocument120 paginiRadiologie Pentru Studenti Si Medici StomatologiAndreea Mirea100% (1)
- Afte Si AftozeDocument10 paginiAfte Si AftozeMadalina SfredelÎncă nu există evaluări
- Grile Med InternaDocument4 paginiGrile Med InternaAndreea MireaÎncă nu există evaluări
- 6 Caria Precoce A CopilarieiDocument9 pagini6 Caria Precoce A CopilarieiAlexandra Nitu100% (1)
- Protezarea ProvizorieDocument47 paginiProtezarea ProvizorieAna Maria100% (2)
- Ciment Are ADocument17 paginiCiment Are AAndreea MireaÎncă nu există evaluări
- Anomalii CromozomialeDocument39 paginiAnomalii CromozomialeAndreea MireaÎncă nu există evaluări
- Organizarea Sistemului ImunDocument69 paginiOrganizarea Sistemului ImunAndreea MireaÎncă nu există evaluări
- Miologia MembrelorDocument8 paginiMiologia MembrelorAndreea MireaÎncă nu există evaluări
- Metode Radio ImagisticeDocument29 paginiMetode Radio ImagisticeAndreea MireaÎncă nu există evaluări
- Of Ta Lomo LogieDocument121 paginiOf Ta Lomo LogieMiruna Alexandra Dima100% (1)
- Nervul TrigemenDocument4 paginiNervul TrigemenAndreea MireaÎncă nu există evaluări
- Caracterizare Paralel MorometiiDocument3 paginiCaracterizare Paralel MorometiiAndreea Mirea100% (1)
- Geografia Sectorului 5Document7 paginiGeografia Sectorului 5Andreea MireaÎncă nu există evaluări
- Materiale DentareDocument83 paginiMateriale DentareAndreea Mirea100% (1)
- MioritaDocument2 paginiMioritaAndreea Mirea100% (1)
- ZburatorulDocument1 paginăZburatorulAndreea MireaÎncă nu există evaluări
- MorometiiDocument1 paginăMorometiiAndreea MireaÎncă nu există evaluări