S-ar putea să vă placă și
- Receptori BiochimieDocument96 paginiReceptori Biochimiedeynyk0% (1)
- Imunitatea InnascutaDocument30 paginiImunitatea InnascutaKfa Dkh100% (1)
- Homeostazia Mediului InternDocument8 paginiHomeostazia Mediului InternMarcu DanielaÎncă nu există evaluări
- Glicoliza PiruvicDocument15 paginiGlicoliza PiruvicTărnăuceanu Cosmin ConstantinÎncă nu există evaluări
- Lipoproteinele PlasmaticeDocument19 paginiLipoproteinele PlasmaticeAncuta CasapuÎncă nu există evaluări
- 5.metabolismul Glucidelor 1Document10 pagini5.metabolismul Glucidelor 1Ioana Estela PătrulescuÎncă nu există evaluări
- Biochimie Clinica Curs 6Document23 paginiBiochimie Clinica Curs 6Karina DeniceÎncă nu există evaluări
- Biosinteza Lipidelor Curs 8Document64 paginiBiosinteza Lipidelor Curs 8Rotaru Lacramioara OanaÎncă nu există evaluări
- Testul PCRDocument9 paginiTestul PCRIulia BrindescuÎncă nu există evaluări
- Biochimie MedicalaDocument133 paginiBiochimie MedicalaPaula AlexandraÎncă nu există evaluări
- Cap.10 Sist - Imunitar - Histologie AnimalaDocument34 paginiCap.10 Sist - Imunitar - Histologie AnimalaraduvascautiÎncă nu există evaluări
- Subiecte Simpificat BiochimieDocument4 paginiSubiecte Simpificat Biochimiemarinescu danÎncă nu există evaluări
- CURS 3 - Structura Celulei BacterieneDocument6 paginiCURS 3 - Structura Celulei BacterienesorinnafÎncă nu există evaluări
- Celula, Proprietatile CeluleiDocument12 paginiCelula, Proprietatile CeluleiAlexÎncă nu există evaluări
- Curs FiziopatologieDocument52 paginiCurs FiziopatologieAna-MariaParaschiv100% (1)
- Curs Nr. 10 Apoptoza Si NecrozaDocument67 paginiCurs Nr. 10 Apoptoza Si NecrozaLarisa LașcuÎncă nu există evaluări
- Homeostazia Metabolica - Cu PaginiDocument167 paginiHomeostazia Metabolica - Cu PaginiCezaraJalencoÎncă nu există evaluări
- Reticulul Endoplasmatic Si RibozomiiDocument6 paginiReticulul Endoplasmatic Si RibozomiiallthefeelingsÎncă nu există evaluări
- Metab. Glucidic - Curs 1, BiochimieDocument46 paginiMetab. Glucidic - Curs 1, BiochimieGreta IulianaÎncă nu există evaluări
- Curs 2 Biochimie Clinica PDFDocument9 paginiCurs 2 Biochimie Clinica PDFCristina MurgociÎncă nu există evaluări
- ElectroforezaDocument12 paginiElectroforezaOana Loredana CoşmanÎncă nu există evaluări
- Bioetica CAP VII Genet Genom OKDocument5 paginiBioetica CAP VII Genet Genom OKBarbu ValentinÎncă nu există evaluări
- CARTE Imunologie 2006Document83 paginiCARTE Imunologie 2006yea, right100% (1)
- Referat Biochimie Compozitia Chimica A SangeluiDocument6 paginiReferat Biochimie Compozitia Chimica A SangeluiGambleru IonutzÎncă nu există evaluări
- Curs 4 - Trombocite, HemostazaDocument14 paginiCurs 4 - Trombocite, HemostazaCiprian Rata100% (1)
- Curs Capitolul VIII Tema 1 2 Neurofiziologie Motorie Și Integrativă - 48797Document171 paginiCurs Capitolul VIII Tema 1 2 Neurofiziologie Motorie Și Integrativă - 48797Catalina EsanuÎncă nu există evaluări
- FitosterideDocument2 paginiFitosterideSelena PufarinÎncă nu există evaluări
- C14.1 GlucocorticoiziiDocument20 paginiC14.1 GlucocorticoiziiGeo GrigoreÎncă nu există evaluări
- Membrane Biologice 1Document10 paginiMembrane Biologice 1Anca NegreanÎncă nu există evaluări
- Metabolismul Intermediar Al GlucidelorDocument20 paginiMetabolismul Intermediar Al GlucidelorCristina GheraÎncă nu există evaluări
- Starea de Sanatate Si Conceptul de BoalaDocument48 paginiStarea de Sanatate Si Conceptul de BoalaRobertÎncă nu există evaluări
- Curs NR 7 HormoniiDocument9 paginiCurs NR 7 HormoniiCCCAXAÎncă nu există evaluări
- Curs 14 Ingineria Genetica Si Biotehnologiile ModerneDocument8 paginiCurs 14 Ingineria Genetica Si Biotehnologiile ModerneDavid SimionÎncă nu există evaluări
- Metabolismul-Lipidic Sinteza ReactiiDocument35 paginiMetabolismul-Lipidic Sinteza ReactiiAna-Maria LupuÎncă nu există evaluări
- Mescalina Referat BunDocument6 paginiMescalina Referat BunSimona SticletÎncă nu există evaluări
- AcetilcolinăDocument4 paginiAcetilcolinăjust_joyÎncă nu există evaluări
- GlicocalixDocument16 paginiGlicocalixivanalexandra0% (1)
- Alergii Si Plante Alergene-xBOOKSDocument0 paginiAlergii Si Plante Alergene-xBOOKSalbudfÎncă nu există evaluări
- Lucrare Disertatie - Lulea Andreea Catalina - 2018, FINALDocument43 paginiLucrare Disertatie - Lulea Andreea Catalina - 2018, FINALAndreea LuleaÎncă nu există evaluări
- CORTICOSTEROIZII Corticosteroizii CursDocument6 paginiCORTICOSTEROIZII Corticosteroizii CursungabiÎncă nu există evaluări
- CELULADocument7 paginiCELULAMadalina FoteaÎncă nu există evaluări
- Glandele SuprarenaleDocument2 paginiGlandele Suprarenaleirinamelnic96Încă nu există evaluări
- Metabolismul LipidicDocument66 paginiMetabolismul LipidicCalina Fodor100% (1)
- Citokine 2019Document36 paginiCitokine 2019Bubulac AngelaÎncă nu există evaluări
- CURS Receptori Biochimie - UMFDocument25 paginiCURS Receptori Biochimie - UMFAlexcsandruÎncă nu există evaluări
- Aspecte Generale Ale Semnalizării CelulareDocument65 paginiAspecte Generale Ale Semnalizării CelulareAlina MihuÎncă nu există evaluări
- 3.semnalizare CelularaDocument15 pagini3.semnalizare CelularaManu El100% (1)
- C12-13semnalizarea CelularăDocument35 paginiC12-13semnalizarea CelularăCezara AxinteÎncă nu există evaluări
- ReceptoriiDocument96 paginiReceptoriiAndreea IvanescuÎncă nu există evaluări
- Semnalizare Intracelulara SintezaDocument24 paginiSemnalizare Intracelulara SintezaBuliga Claudia100% (1)
- Semnalizarea CelularaDocument71 paginiSemnalizarea CelularaOvidiu VaidaÎncă nu există evaluări
- Subiect 9Document5 paginiSubiect 9DAN-ANDREI DARIEÎncă nu există evaluări
- SNVDocument30 paginiSNVEugenia MirosnicencoÎncă nu există evaluări
- Receptor IDocument96 paginiReceptor IDan RuicanÎncă nu există evaluări
- Curs 13 BIOCELDocument14 paginiCurs 13 BIOCEL12052000Încă nu există evaluări
- SemnalizareDocument3 paginiSemnalizareSuiu-Tincu ClaudiaÎncă nu există evaluări
- Fiziologie AnimalaDocument25 paginiFiziologie AnimalafasolăiÎncă nu există evaluări
- Curs Sem I - An IV - Farmacologie Si Farmacoterapie PDFDocument372 paginiCurs Sem I - An IV - Farmacologie Si Farmacoterapie PDFAndreea SanduÎncă nu există evaluări
- Notiuni in NeurofiziologieDocument13 paginiNotiuni in NeurofiziologieAlexandra NistorÎncă nu există evaluări
- Bazele Neurochimice Ale Comportamentului TotDocument13 paginiBazele Neurochimice Ale Comportamentului TotDiana BontoÎncă nu există evaluări
- LegalitateDocument1 paginăLegalitatevlad_văluÎncă nu există evaluări
- Biochimie - Metabolismul GeneralitatiDocument10 paginiBiochimie - Metabolismul Generalitativlad_văluÎncă nu există evaluări
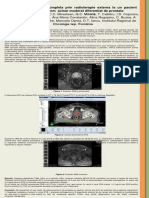
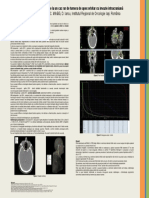
- Regresie Completa Prin Radioterapie Externa La Un Pacient Cu Adenocarcinom de ProstataDocument2 paginiRegresie Completa Prin Radioterapie Externa La Un Pacient Cu Adenocarcinom de Prostatavlad_văluÎncă nu există evaluări
- Regresie Completa Prin Radioterapie Externa La Un Pacient Cu Adenocarcinom de ProstataDocument1 paginăRegresie Completa Prin Radioterapie Externa La Un Pacient Cu Adenocarcinom de Prostatavlad_văluÎncă nu există evaluări
- Drepturi Salariale OctombrieDocument8 paginiDrepturi Salariale Octombrievlad_văluÎncă nu există evaluări
- GammaMedplus IX Instructions For Use 2011-11-15 - RODocument302 paginiGammaMedplus IX Instructions For Use 2011-11-15 - ROvlad_văluÎncă nu există evaluări
- Regresie Completa Prin Radioterapie Externa La Un Pacient Cu Adenocarcinom de ProstataDocument2 paginiRegresie Completa Prin Radioterapie Externa La Un Pacient Cu Adenocarcinom de Prostatavlad_văluÎncă nu există evaluări
- Poster SinaiaDocument1 paginăPoster Sinaiavlad_văluÎncă nu există evaluări
- 2011mardanet A.F. Analiza Instrumentala Partea I Cap. 1.1 - 1.9Document110 pagini2011mardanet A.F. Analiza Instrumentala Partea I Cap. 1.1 - 1.9ionut100% (3)
- Reviste StiintificeDocument1 paginăReviste Stiintificevlad_văluÎncă nu există evaluări
- Poster MoinestiDocument1 paginăPoster Moinestivlad_văluÎncă nu există evaluări
- Examen Asistent Cercetare - StefanestiDocument13 paginiExamen Asistent Cercetare - Stefanestivlad_văluÎncă nu există evaluări
- Analiza Instrumentala SpectometrieDocument7 paginiAnaliza Instrumentala Spectometrievlad_văluÎncă nu există evaluări
- 12 Mai, Ziua Internațională A Asistentului Medical: Specializarea Asistent Medical GeneralistDocument4 pagini12 Mai, Ziua Internațională A Asistentului Medical: Specializarea Asistent Medical Generalistvlad_văluÎncă nu există evaluări
- MCE2 MG TimpuluiDocument35 paginiMCE2 MG TimpuluiLaura CasandriucÎncă nu există evaluări
- Metode Didactice InteractiveDocument7 paginiMetode Didactice Interactivevlad_văluÎncă nu există evaluări
- Eco EtoDocument1 paginăEco Etovlad_văluÎncă nu există evaluări
- Lucrari Practice EntomologieDocument1 paginăLucrari Practice EntomologieAlexandraPricopeÎncă nu există evaluări
- Receptori Nucleari OrfaniDocument12 paginiReceptori Nucleari Orfanivlad_văluÎncă nu există evaluări
- Specii IerbarDocument2 paginiSpecii Ierbarvlad_văluÎncă nu există evaluări
- Ecologie Acvatica - BrezeanuDocument76 paginiEcologie Acvatica - Brezeanudaniral100% (1)
- CR 10.1 Cerinte Specifice 30.10.2012 PDFDocument13 paginiCR 10.1 Cerinte Specifice 30.10.2012 PDFGeorge Mihail ManÎncă nu există evaluări
- MCE6 - Disciplina ScolaraDocument74 paginiMCE6 - Disciplina ScolaraMoraruElenaÎncă nu există evaluări
- Comunicarea Celulara NouDocument20 paginiComunicarea Celulara Nouvlad_văluÎncă nu există evaluări
- Citofiziologie VegetalaDocument23 paginiCitofiziologie Vegetalavlad_văluÎncă nu există evaluări
- Fanerogame Parazite Si Plante CarnivoreDocument25 paginiFanerogame Parazite Si Plante Carnivorevlad_văluÎncă nu există evaluări
- Cresterea Si Dezvoltarea La PlanteDocument16 paginiCresterea Si Dezvoltarea La Plantevlad_văluÎncă nu există evaluări
- Regimul de Apa Al PlantelorDocument20 paginiRegimul de Apa Al PlantelorStefan100% (3)
- Respiratia Plantelor 2015Document14 paginiRespiratia Plantelor 2015StefanÎncă nu există evaluări