S-ar putea să vă placă și
- Curs BMCDocument106 paginiCurs BMCElena Cojocaru100% (1)
- HomeopatieDocument62 paginiHomeopatieElena CojocaruÎncă nu există evaluări
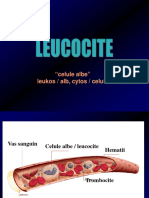
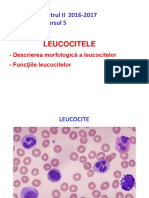
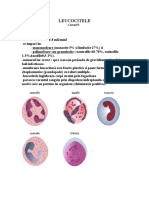
- LeucociteleDocument23 paginiLeucociteleAna-Maria Ene100% (2)
- Curs 5 Si 6 Leucocite Si Trombocite PowerPoint PresentationDocument113 paginiCurs 5 Si 6 Leucocite Si Trombocite PowerPoint PresentationAdrian-Cristi PredaÎncă nu există evaluări
- Imunitatea CelularaDocument6 paginiImunitatea Celulara11224499100% (1)
- Leucocite Si TrombociteDocument113 paginiLeucocite Si TrombociteDana Chitoi100% (1)
- Grile OrtoDocument3 paginiGrile OrtoElena CojocaruÎncă nu există evaluări
- CitokineleDocument3 paginiCitokineleatezorÎncă nu există evaluări
- Curs 4 PDFDocument55 paginiCurs 4 PDFladyddyÎncă nu există evaluări
- Curs 3 - LeucocitulDocument21 paginiCurs 3 - LeucocitulAndreeaAllexandraÎncă nu există evaluări
- InflamatiaDocument25 paginiInflamatiaPELERINUL78Încă nu există evaluări
- ImunologieDocument10 paginiImunologiexsmileÎncă nu există evaluări
- Fiziologie (Cursul 5)Document27 paginiFiziologie (Cursul 5)laviniapatrascoiuÎncă nu există evaluări
- Curs 3 - LeucocitulDocument21 paginiCurs 3 - LeucocitulNihilobstatÎncă nu există evaluări
- Leuco Cite LeDocument5 paginiLeuco Cite LeciprianÎncă nu există evaluări
- InflamatiaDocument5 paginiInflamatiaComan Bogdan100% (1)
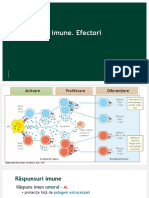
- Raspuns Imun UmoralDocument9 paginiRaspuns Imun UmoralAmelia NegoiÎncă nu există evaluări
- Citokine 2019Document36 paginiCitokine 2019Bubulac AngelaÎncă nu există evaluări
- Citokinele: Diferente Fata de Hormoni: Hormonii Clasici Circula in Concentratii Nanomolare, in Timp Ce Unele CitokineDocument5 paginiCitokinele: Diferente Fata de Hormoni: Hormonii Clasici Circula in Concentratii Nanomolare, in Timp Ce Unele CitokineIulia DragomirÎncă nu există evaluări
- CURS 4 (C, IFN, Citokine, Chemokine)Document47 paginiCURS 4 (C, IFN, Citokine, Chemokine)Cristian Rusu100% (1)
- Imunologie Curs 5 APCDocument4 paginiImunologie Curs 5 APCRaluca MosoiaÎncă nu există evaluări
- Procesează Prezintă: Exo T Helper CD4+Document3 paginiProcesează Prezintă: Exo T Helper CD4+Alex StefanÎncă nu există evaluări
- Produșii de Secreție Ai Celulelor Activate În RIUDocument6 paginiProdușii de Secreție Ai Celulelor Activate În RIUClaudia SmaduÎncă nu există evaluări
- Imunologie LPDocument9 paginiImunologie LPLUÎncă nu există evaluări
- Curs 5 RI - Part 5Document18 paginiCurs 5 RI - Part 5AuraÎncă nu există evaluări
- Lim Fokine LeDocument8 paginiLim Fokine LePatricia TamasÎncă nu există evaluări
- (2023) .Limfocitele T.Răspuns CelularDocument48 pagini(2023) .Limfocitele T.Răspuns CelularTanea MoiseiÎncă nu există evaluări
- Curs InflamatieDocument10 paginiCurs Inflamatiebogdanw11Încă nu există evaluări
- Curs 6 Fiziologie An II Ro Sem II Seriile Abcd 2021Document61 paginiCurs 6 Fiziologie An II Ro Sem II Seriile Abcd 2021Cristina THCSÎncă nu există evaluări
- InflamatiiDocument43 paginiInflamatiiW205 SamÎncă nu există evaluări
- Imunopat Curs 3Document14 paginiImunopat Curs 3Ioana VoinescuÎncă nu există evaluări
- CITOKINEDocument11 paginiCITOKINENdbebe BebeÎncă nu există evaluări
- Curs 4Document7 paginiCurs 4Dorin SaulescuÎncă nu există evaluări
- Curs 6Document7 paginiCurs 6PopescuÎncă nu există evaluări
- GranulopoiezaDocument34 paginiGranulopoiezaElly ElizaÎncă nu există evaluări
- Imuno 2 Dr. C. StoiaDocument24 paginiImuno 2 Dr. C. StoiaAdelleBordeaÎncă nu există evaluări
- HemostazaDocument23 paginiHemostazaLarisa DumitruÎncă nu există evaluări
- Sist Circ - 2014Document40 paginiSist Circ - 2014Anton PannÎncă nu există evaluări
- Curs 7 Petru OntogenieDocument11 paginiCurs 7 Petru OntogenieAd AdrianaÎncă nu există evaluări
- Curs6 MD RODocument13 paginiCurs6 MD ROStef DavidÎncă nu există evaluări
- Mediatori ImuniDocument56 paginiMediatori ImuniBogdan Adriana100% (1)
- Diferențiere Fenotipică Și Tipuri LT. NK Și NKTDocument5 paginiDiferențiere Fenotipică Și Tipuri LT. NK Și NKTClaudia SmaduÎncă nu există evaluări
- Curs 8 FiziologieDocument4 paginiCurs 8 Fiziologiedoll_andaÎncă nu există evaluări
- Interleukin 1Document14 paginiInterleukin 1Dragoș MălăescuÎncă nu există evaluări
- Curs 3 Organizarea Sistemului ImunDocument5 paginiCurs 3 Organizarea Sistemului ImunAdaÎncă nu există evaluări
- BazofilulDocument29 paginiBazofilulCristina Tomescu100% (1)
- Imuno-Cooperare CelularaDocument8 paginiImuno-Cooperare CelularaBianca BradÎncă nu există evaluări
- Curs 2Document54 paginiCurs 2Bianca Gabriela ZanfirÎncă nu există evaluări
- Fiziopatologia Curs Inflamatie. DR NemesDocument10 paginiFiziopatologia Curs Inflamatie. DR NemesTony TudorÎncă nu există evaluări
- LeucociteDocument23 paginiLeucociteAna-Maria EneÎncă nu există evaluări
- Rezumat ImunologieDocument23 paginiRezumat ImunologieDorinaBunciucÎncă nu există evaluări
- Curs 7 Imunitate 2Document75 paginiCurs 7 Imunitate 2Daniela PopaÎncă nu există evaluări
- Imuno Curs1Document6 paginiImuno Curs1cnajesrnasiferfÎncă nu există evaluări
- Curs 2 Concepte Referitoare La Apararea AntiinfectioasaDocument51 paginiCurs 2 Concepte Referitoare La Apararea AntiinfectioasatimcarasÎncă nu există evaluări
- C3. Leucocitele Si TrombociteleDocument63 paginiC3. Leucocitele Si TrombociteleMirabelaRÎncă nu există evaluări
- C01. Imunitatea InnascutaDocument8 paginiC01. Imunitatea InnascutaAlin AllinÎncă nu există evaluări
- FIZIOLOGIE Sangele2Document63 paginiFIZIOLOGIE Sangele2Mihaela VadanaÎncă nu există evaluări
- Histologie Curs 05Document9 paginiHistologie Curs 05Carmen-Elena ManeaÎncă nu există evaluări
- Curs 10Document68 paginiCurs 10Teo TeoÎncă nu există evaluări
- Metabolismul BacteriilorDocument12 paginiMetabolismul BacteriilorElena CojocaruÎncă nu există evaluări
- Curs 3 - CirculatiaDocument54 paginiCurs 3 - CirculatiaElena CojocaruÎncă nu există evaluări
- Curs 4 - Circulatia 2Document70 paginiCurs 4 - Circulatia 2Brigitte Si AlinÎncă nu există evaluări
- MED BG 1bDocument18 paginiMED BG 1bAna IlieÎncă nu există evaluări
- Istoricul Microbiologiei - I: - Aristotle Teorie Idealist Ă Teorii Materialiste AnticeDocument19 paginiIstoricul Microbiologiei - I: - Aristotle Teorie Idealist Ă Teorii Materialiste AnticeElena CojocaruÎncă nu există evaluări
- Curs 2 EpidemiologieDocument20 paginiCurs 2 EpidemiologieElena CojocaruÎncă nu există evaluări
- Curs EpidemiologieDocument16 paginiCurs EpidemiologieElena CojocaruÎncă nu există evaluări
- Curs Introducere 2018 PneumologieDocument5 paginiCurs Introducere 2018 PneumologieElena CojocaruÎncă nu există evaluări
- SPM Cursuri 2020Document176 paginiSPM Cursuri 2020Elena CojocaruÎncă nu există evaluări
- OncologieDocument112 paginiOncologieElena Cojocaru0% (1)
- Cursuri Boli InfectioaseDocument162 paginiCursuri Boli InfectioaseElena CojocaruÎncă nu există evaluări
- Istoria Naturala 2018 PneumologieDocument4 paginiIstoria Naturala 2018 PneumologieElena CojocaruÎncă nu există evaluări
- Grile OrtopedieDocument2 paginiGrile OrtopedieElena CojocaruÎncă nu există evaluări
- Ghid ESCDocument46 paginiGhid ESCElena CojocaruÎncă nu există evaluări
- Curs 2Document1 paginăCurs 2Elena CojocaruÎncă nu există evaluări
- Grile OrtopedieDocument2 paginiGrile OrtopedieElena CojocaruÎncă nu există evaluări
- Curs Fizpat 7 1Document4 paginiCurs Fizpat 7 1Elena CojocaruÎncă nu există evaluări
- Fiziopatologie Curs 5 1Document2 paginiFiziopatologie Curs 5 1Elena CojocaruÎncă nu există evaluări
- Fiziopatologie Curs 10 1Document5 paginiFiziopatologie Curs 10 1Elena CojocaruÎncă nu există evaluări
- Curs 4Document3 paginiCurs 4Elena CojocaruÎncă nu există evaluări
- Tulburari CirculatoriiDocument22 paginiTulburari CirculatoriiElena CojocaruÎncă nu există evaluări
- NEOPLAZIADocument30 paginiNEOPLAZIAElena CojocaruÎncă nu există evaluări
- IMUNOPATOLOGIEDocument15 paginiIMUNOPATOLOGIEElena Cojocaru0% (1)
- 03 Ok INFLAMATIADocument43 pagini03 Ok INFLAMATIAali_hgyÎncă nu există evaluări
- Capitolul IIIDocument25 paginiCapitolul IIIElena CojocaruÎncă nu există evaluări
- Capitolul IIDocument28 paginiCapitolul IIElena CojocaruÎncă nu există evaluări
- Patologie CelularaDocument25 paginiPatologie CelularaElena CojocaruÎncă nu există evaluări